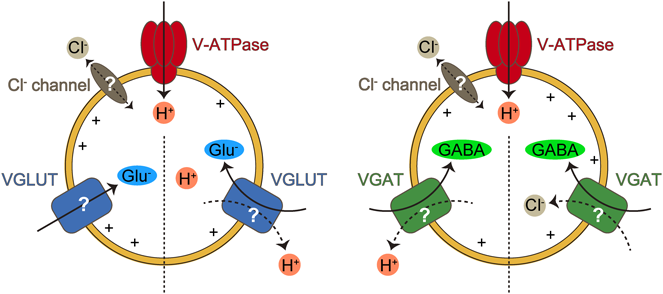
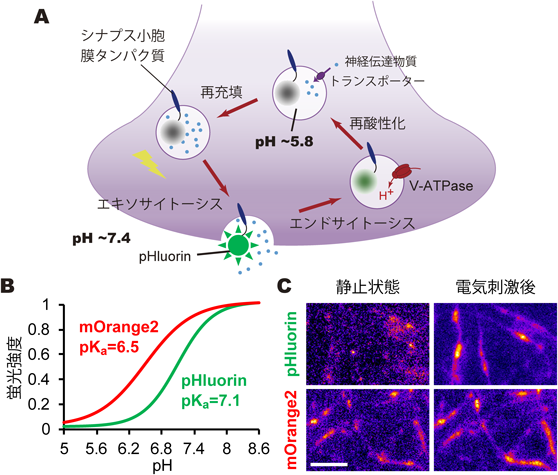
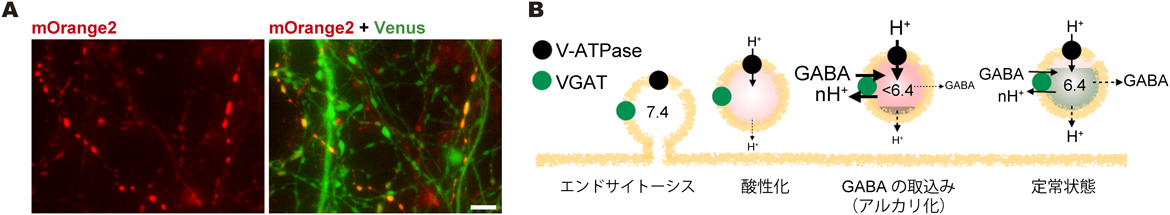
神経伝達物質のシナプス小胞内輸送機構Mechanism of vesicular neurotransmitter transport into the synaptic vesicles
1 大阪医科大学医学部生命科学講座生理学教室Osaka Medical College, Faculty of Medicine, Division of Life Sciences, Department of Physiology ◇ 〒569–8686 大阪府高槻市大学町2–7 ◇ 2–7, Daigaku-machi, Takatsuki, Osaka, 569–8686, Japan
2 同志社大学大学院脳科学研究科神経膜分子機能部門Doshisha University, Graduate school of Brain Science, Laboratory of Neural Membrane Biology ◇ 〒610–0394 京都府京田辺市多田羅都谷1–3 ◇ 1–3, Tatara-Miyakodani, Kyotanabe, Kyoto, 610–0394, Japan
発行日:2017年12月25日Published: December 25, 2017