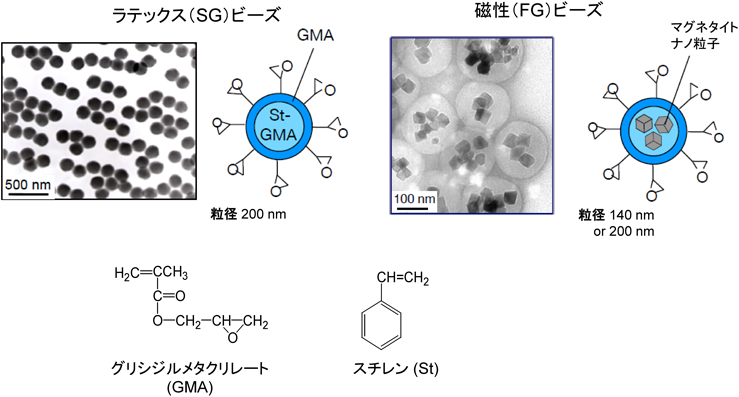
私は,ボストンにあるマサチュセッツ工科大学(MIT)のP. Sharp教授(1993年分断遺伝子の発見でノーベル医学生理学賞受賞)に師事し,遺伝子発現制御という分子生物学における世界最先端の研究にじかにふれることができた.その貴重な留学時代の経験に基づいて,「サイエンスは個性とプライド」と「独自の技術開発は独創的研究を生む」の二つを研究モットーとして,学際的研究を積極的に取り入れて新たな技術開発に努めてきた.その結果,半田ビーズと呼ばれるナノサイズのラテックス(SG)ビーズおよび磁性(FG)ビーズを開発し(図1),それを用いた革新的なアフィニティ精製技術を確立することができた1–3).
目的タンパク質を単離・精製するために,これまで使用されてきたカラムクロマト法は,分離原理に基づいて疎水的,静電的,アフィニティ,ゲルろ過に大別されるが,いずれのカラムクロマト法の回収効率もおよそ10%以下しかなく,105種以上のタンパク質ライブラリーから1種の目的タンパク質を精製するには,異なるカラムクロマト法を五つ以上組み合わせた多工程が必要になる.したがって,最終的な目的タンパク質の精製・純化に到達するまでには,多量の出発材料,時間,労力,経費,ノウハウを要し,しかも,各工程における目的タンパク質の損失,失活,分解は必至で,多工程になるとそれらがさらに加算される.したがって,従来のカラムクロマト法では,薬剤ターゲットなどの目的タンパク質を単離・純化することは至難の業であった.
私は,目的タンパク質を単離・純化する際の重要な課題は,①精製物の純度(purity)を高め,②回収率(recovery rate)を向上し,③活性(activity)を維持することと考えた.それを達成するためには,工程数を減らし,非特異的結合タンパク質の混入を減らし,回収率を高める工夫が必要となる.そこでまず,工程数を減らすためにアフィニティ精製を前提として,従来のカラムクロマト法の主な欠点がアガロース担体とカラム法に起因すると考え,その解決策を入念に練った.その結果,これまで金科玉条のごとく使われてきたマイクロ(µ)サイズのアガロース担体に取って代わるナノ(n)サイズの新規ビーズ担体を独自に作製し,カラム法に代わるバッチ法でアフィニティ精製することを考えた.新規ビーズ担体を作製するのに,高分子化学および磁気工学の専門家である川口春馬名誉教授(慶大・理工)および阿部正紀名誉教授(東工大・理工)と共同研究を行って,専門の知識や技術を学びながら,まず,ラテックス(SG)ビーズの作製を猪俣幸雄博士(現花王株式会社)が成功させ,次に,磁性体であるマグネタイトをポリマーで被覆した磁性(FG)ビーズの作製を西尾広介博士(現テルモ株式会社)が成功させた(図1).これらナノサイズの新規ビーズ担体表面に各種リガンドを官能基を介して固定化し,それをタンパク質ライブラリーと混合して,バッチ法による単一工程でのアフィニティ精製技術の確立を目指した.各反応条件を入念に検討し,いろいろな試行錯誤を繰り返しながら,当初の目的を何とか達成することができた1–3).
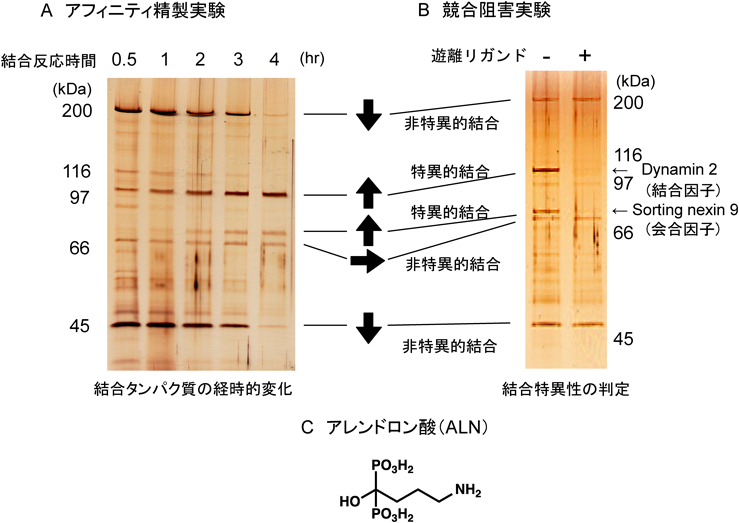
まず,作製した新規ビーズ担体は,非特異的タンパク質の結合がきわめて少ない優れた表面性状を持ち,粒径がナノサイズなので単位容積あたりの粒子総表面積を大きく確保できる.また,表面を構成するグリシジルメタクリレート(glycidyl methacrylate:GMA)は反応性官能基であるエポキシ基(107個以上)を持つので,多量のリガンドをエポキシ基を介して粒子表面に固定化できる.その結果,精製物の回収率および純度をいずれも想定以上に高めることに成功した.また,ナノサイズのビーズ担体は高分散性・可動性で,バッチ法で行う限り,タンパク質がビーズ上の固定化リガンドといったん結合した後に解離しても,近接するたくさんの固定化リガンドとすぐに再結合および再々結合することが可能になる.したがって,結合反応の開始時はビーズ上の固定化リガンドに,疎水的結合や静電的結合によって低親和性の非特異的タンパク質が多く結合しているが,反応時間が長くなるにつれて,固定化リガンドと特異的かつより強く結合するタンパク質が次第に非特異的結合タンパク質と置き換わってビーズ上に増加する.その結果,結合反応が数時間以上に伸びると,固定化リガンドと特異的にしかもより強く結合するタンパク質がビーズ上に優先的に増えることから,当然,ターゲットを含む特異的結合タンパク質が高回収率かつ高純度に単離・純化できることになる(図2).加えて,ビーズ担体は有機溶媒に耐性なので,難水溶性の有機溶媒にしか溶けないリガンドでも有機溶媒中でビーズ表面に多量に固定化できる.また,そのリガンド固定化ビーズは,水溶液中でリガンドと結合するタンパク質の単離・純化に使用できるので,その汎用性を高めることができた3, 4).
実際に,薬剤を固定化したSG/FGビーズを用いると,数十万種のタンパク質を含む細胞・組織破砕液(タンパク質ライブラリー)からワンステップで,薬剤と結合するタンパク質やタンパク質複合体を高回収率かつ高純度に単離・純化できる.加えて,このシステムでは結合の特異性や強さを容易に判定できるので,薬剤結合タンパク質の中からターゲット候補を推測することができる.このビーズ技術は,従来のカラム法では到底できなかったことを実現可能にしたことで,国の内外を問わず高く評価され,需要もあることから実用化に至っている.ちなみに,アフィニティ精製用のSG/FGビーズは2009年から販売が開始されている.
また,SG/FGビーズは,薬剤の他に各種の代謝産物,有害物質,核酸,タンパク質/ペプチドなどと特異的に結合するタンパク質を単離・同定することができるので,幅広い分野で,しかも基礎研究ならびに応用研究のいずれにおいても使用されている.
近年,FGビーズにより,これまで正確な定量が困難であった血中エキソソームの定量測定が可能になった.簡単に紹介すると,エキソソームと反応する一次抗体を固定化したFGビーズと,二次抗体を固定化した260 nmサイズ溝を持つ光ディスク(DVD)とを用いて,サイズ制限下でエキソソームをサンドイッチすることで,ディスク溝上の二次抗体と反応したエキソソームと1:1で結合するFGビーズの数を高速でカウントする方法である5).抗体固定化FGビーズとともに「ExoCounter」と名づけられた測定装置がJVCケンウッド株式会社で開発され,シスメックス株式会社から販売されている.この装置を用いて,エキソソーム数を測定した結果,乳がんおよび卵巣がんの患者でHer2陽性のエキソソームが有意に増加していることがわかり,現在,その詳細を解析するとともに,他のがんへの有効性が検討されている.
それに加えて,近年,酸化鉄マグネタイトと一緒に蛍光化合物を内包した蛍光磁性(FF)ビーズを開発することに成功した6).抗体固定化FFビーズは,磁石による磁気捕集・濃縮により特異的結合反応を迅速化でき,これまで数十分から数時間以上かかった抗原抗体反応を,わずか1分以内に短縮可能になった.また,磁石を用いた磁気洗浄により非特異的結合FFビーズを1分以内に剥離でき,それによりバックグラウンドノイズを著しく低減できるようになった.この磁気捕集と磁気洗浄により抗原抗体反応時間を驚異的に短縮することが可能になった.さらに,ビーズ中に106個以上の蛍光化合物を内包しているので酵素反応やPCRなどによるシグナル増幅反応が不要で,抗原抗体反応後に直ちに蛍光検出できるという長所を持っている.したがって,抗原抗体反応に基づく測定系であるサンドイッチイムノアッセイや免疫組織染色など,これまでは少なくとも数十分から数時間かかっていたものが,3分以内ときわめて迅速に検出することが可能になった6).現在,それらアッセイ系の自動化装置の開発が試みられている.このFFビーズによる迅速高感度診断システムは,手術中,診察中,救急時,空港検疫時などの迅速・高感度な確定診断を可能にし,患者のQOLの向上,生命の維持,疾病への迅速・的確な対応,最適な手術法の選択などに大いに役立つことが期待される.
1)転写因子ファミリーに属する全メンバーのワンステップ精製
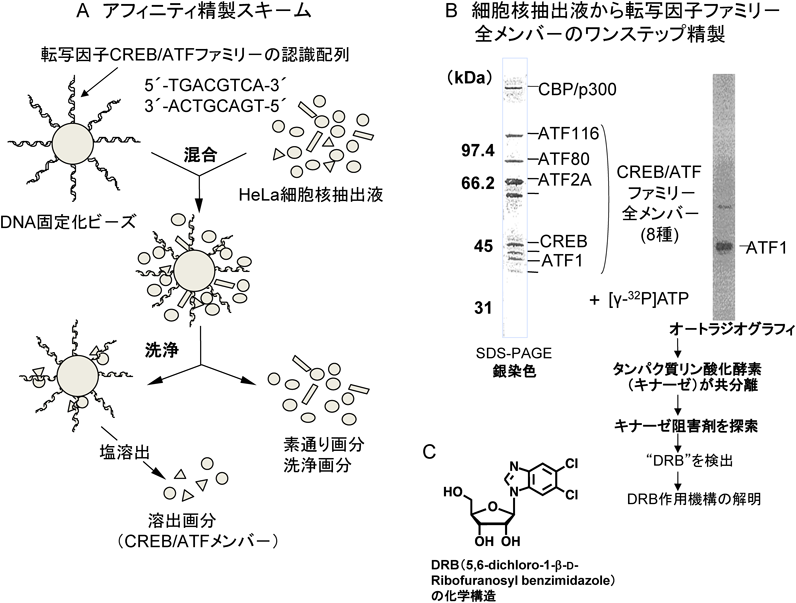
まず,転写因子CREB/ATFファミリーに属するすべてのメンバーが,SG/FGビーズを用いたアフィニティビーズ技術でどのように精製できるかを試みた.CREB/ATFファミリーの認識・結合配列は5′-TGA CGT CA-3′である.そこで,その5′末端にCCC CC(5×C)配列をつけた一本鎖(ss)DNAと,その特異結合配列に相補的な塩基配列の3′末端にGGG GG(5×G)配列をつけたssDNAを合成し,適当な条件下で両者をハイブリッドすると,両末端に5×C配列と5×G配列の一本鎖突出末端を持つ二本鎖(ds)DNAができる.それをアニーリングすると,末端の5×C配列と5×G配列がハイブリッドを形成して平均長が150~250塩基対からなるdsDNAが形成される.その5′末端には5×C, 3′末端には5×Gからなる一本鎖突出部分がある.そこで,そのdsDNAとSG/FGビーズを純水中で混合すると,dsDNAの片末端にある5×G配列のアミノ基とSG/FGビーズ表面のエポキシ基との間で化学結合が起きる.G塩基のアミノ基がエポキシ基との反応性が最も高いことと,dsDNA部分にあるG塩基は水素結合に関与しており,エポキシ基との反応性が弱いことから,5×Gからなる片末端を介してdsDNAをビーズ表面に固定化できる.5×G配列の中にある二つ以上のG塩基が化学結合すると,dsDNAはより強く・安定にビーズ表面に固定化される(図3A).
この特異的塩基配列を10個以上タンデムに持つdsDNAを固定化したSG/FGビーズと培養細胞の核抽出液とを混合し,特異的塩基配列に結合する転写因子のアフィニティ精製を試みた.驚いたことに,CREB/ATFファミリーに属する八つのメンバーがすべて一挙に分離されることがわかった.SDS-PAGEで八つのタンパク質バンドとして分離した後に,各タンパク質をゲルから溶出後に変性・再生して特異的塩基配列への結合能を調べてみると,八つすべてが特異的塩基配列と結合する活性を有することがわかり,CREB/ATF転写因子ファミリーであることが確認できた7, 8).それに加えて,CBP/p300といったコアクチベターやキナーゼ活性が共分離されることもわかった(図3B).このようなことは,従来のDNAアフィニティーカラムクロマト法では到底できなかったので,アフィニティビーズ技術の優位性が証明できた.また,このキナーゼ活性が共分離されたことに興味を持ち,キナーゼ活性の単離・精製を考えたが,同定可能な十分量を得ることが困難だとわかり,まず,このキナーゼ活性の阻害剤を探索してみた.すると,幸運にもアデノシン類似体のDRBを探し当てることができた(図3B).
2)DRBの転写伸長阻害作用
当時,DRBは転写伸長反応を阻害することが報告されており,細胞をDRBで処理すると数十塩基長の短鎖mRNAが蓄積することが知られていた.また,それとは別に,DRBは転写伸長促進活性のあるキナーゼP-TEFb(positive transcription elongation factor b)の阻害剤であることが報告されていた.しかし,DRBの転写伸長阻害メカニズムはまったく不明で,転写伸長阻害とキナーゼP-TEFb活性阻害との関係についても皆目見当がつかなかった.当時は,転写といえば開始反応の制御に皆の興味が注がれていた.転写開始制御に関して,プロモーター上におけるRNAポリメラーゼII(Pol II)と基本転写因子からなる転写開始前複合体形成や,転写開始反応に及ぼすエンハンサーやアクチベーターの作用,コアクチベーターの役割などが次第に解き明かされていたが,転写伸長制御に関しては手のつけどころがなく,皆目わかっていなかった.そんなときに,DNA固定化ビーズによるアフィニティ精製の結果から派生して,DRBと遭遇することができたので,まず,DRBの転写伸長阻害作用とそのメカニズムを自分なりに理解しようと考えた.そこで,MIT時代に修得したin vitro転写系とその再構成系を使って,DRBの作用機構の解析に着手した.まず,プロモーターを含むプラスミドDNAと細胞核粗抽出液によるPol IIと放射能標識した核酸前駆体を混合した転写反応液にDRBを添加すると,フルサイズの転写産物の合成は抑制され,30~50塩基長の短鎖mRNAが蓄積されることがわかった.このin vitro転写系でのDRBによる転写伸長阻害により生じる短鎖mRNAは,すでに報告されていた培養細胞におけるDRB処理時に蓄積する短鎖mRNAと同じ現象であることがわかった.そこで,核粗抽出液をホスホセルロース(P11)カラムを用いて,KCl濃度を0.05 M, 0.3 M, 1.0 Mと段階的に上げて分画し,各濃度における溶出画分の転写活性能を調べた.すると,1.0 M画分にPol IIと基本転写因子のほとんどすべてが溶出し,この画分だけでも細胞核粗抽出液と同様にフルサイズの転写産物が合成された.ところが,1.0 M画分の転写はDRBに耐性で,転写伸長反応は阻害されなかった.しかし,1.0 M画分に0.3 M画分を加えると,DRBによる伸長阻害が起り,短鎖mRNAが産生されることがわかった.この結果は,0.3 M画分にDRB感受性を誘導する活性が存在することを示している.そこで,0.3 M画分をさらに別のカラムクロマト法で分画し,DRB感受性を誘導する活性を持つ画分を見いだし,それをさらに別のカラムクロマト法で分画し,同様のやり方でDRB感受性を誘導する活性を見つけるという手順を繰り返し,七つの異なるカラムクロマト法を組み合わせて,最終的にDRB感受性を誘導する因子として,160 kDaと14 kDaの二つのタンパク質からなる複合体を純化することに成功した.活性には二つの因子がともに必要で,両因子からなる複合体をDSIF(DRB sensitivity-inducing factor)と名づけた9).質量分析によりDSIFは,酵母の遺伝学的解析から同定され機能未知であった転写制御因子Spt5とSpt4のヒトホモログであることがわかった.DSIFの単離・同定は,現大阪大学客員教授の和田忠士博士が行い,現第一三共製薬に勤務する高木敏行博士が積極的に加勢してくれた.また,その当時の生化学に必須な技術であるカラムクロマト法を私のラボに確立し,ラボの皆に啓発してくれたのが現大阪大学大学院工学研究科教授の渡辺肇博士で,ここに心から感謝の意を表したい.
さらに,はじめの細胞粗核抽出液を分画したP11カラムの1.0 M画分を他のカラムクロマトで分画し,Pol IIをはじめ基本転写因子群を高度に精製し,それら高度精製因子群によるin vitro転写再構成系でDRBによる転写伸長阻害効果を検討した.ところが,DRBによる転写伸長阻害はDSIFだけでは起こらず,DSIFに加えて別の因子が必要であることがわかった.その因子を高度精製因子群による再構成系を用いて精製することに成功し,NELF(negative elongation factor)と名づけた10).NELFの単離・精製は,現東京工業大学教授の山口雄輝博士が一人ですべて行った.面白いことに,NELFは下等真核生物である酵母にはなく高等真核生物にのみ存在することがわかり,四つのサブユニットからなる複雑な複合体を,組換えタンパク質を用いて再構成することにも成功した11).
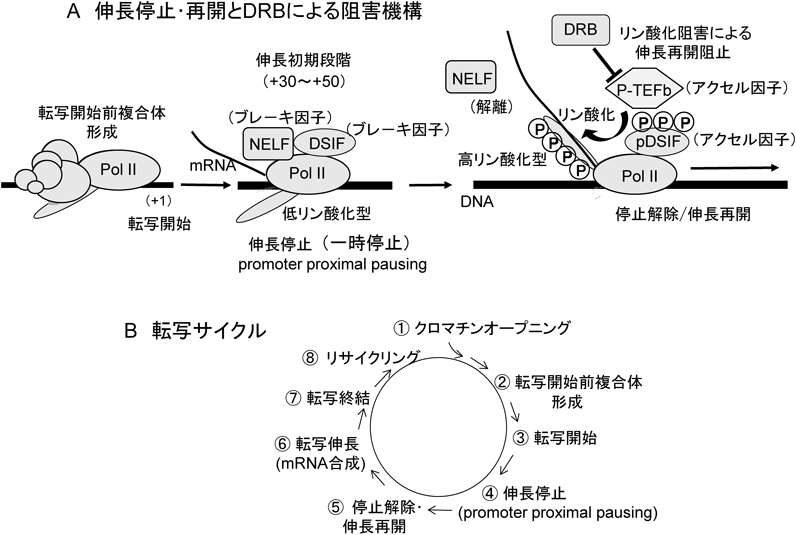
DSIFとNELFは低リン酸化型Pol IIと結合し,プロモーターの近位(+30~+50付近)でPol IIの一時的なmRNA合成停止(伸長停止)を引き起こす.その後,このDSIFとNELFによる伸長停止は,John Lis(コーネル大学)らが熱ショック遺伝子で報告していたPol IIのプロモーター近位停止(promoter proximal pausing)そのものであることがわかった(図4A)12).
次に,この転写伸長段階の一時停止を解除して,Pol IIの伸長再開を引き起こすメカニズムを明らかにすることにも成功した(図4A)13).それに関わる重要な因子がDavid Price(アイオワ大学)らが単離したDRB感受性キナーゼP-TEFbである.P-TEFbはCyclin TとCDK9からなる複合体で,CDK9がキナーゼ活性を持っている.真核生物と原核生物のRNAポリメラーゼの最も大きな違いは,最大サブユニットのC末端ドメイン(C-terminal domain:CTD)にある七つのアミノ酸配列の繰り返し配列で,この配列は原核細胞ではみられない.ヒトは52回の繰り返しを持ち,転写が活発に行われているときのCTDは高リン酸化型で,転写開始前複合体や伸長段階で停止しているときなどでは低リン酸化型を示す.P-TEFbはPol II CTDの七つのアミノ酸配列の2番目のSerをリン酸化する.また面白いことに,P-TEFbはDSIFの160 kDaサブユニットの繰り返し配列(C-terminal region:CTR)もリン酸化する14).P-TEFbによるリン酸化によって,NELFが伸長停止複合体からキックアウトされて解離し,伸長停止が解除されて転写が再開し,伸長反応が転写終結まで継続する.また,DSIFは,NELFとともにPol IIに結合して転写伸長反応を停止するブレーキ因子だが,P-TEFbによってリン酸化されるとアクセル因子に変換する.DSIFがリン酸化されないと,mRNAは200~300塩基長しか合成されないが,P-TEFbによってDSIFがリン酸化されると,数10万塩基長以上の長鎖mRNAが合成される14).その理由として,リン酸化pDSIFには非リン酸化DSIFとは異なるクロマチンリモデリング因子がリクルートされると考えられる.
最後に,DRBの作用機構に関して述べると,DRBはP-TEFbと結合して,P-TEFbによる伸長停止を解除するリン酸化活性を阻害する.したがって,DRBが存在するとPol IIは伸長停止の状態から解除されず,いつまでも伸長停止状態を続けることになる.
我々はPol IIによる転写伸長反応のブレーキ因子としてDSIFとNFLFを単離・同定し,それら因子によるPol IIの伸長停止機構およびP-TEFbによる停止解除・再開機構を明らかにし,加えて,DRBの作用機構を解き明かすことに成功した(図4A)9, 10, 13).2018年に,Max Planck研究所のPatrick Cramer博士らによるPol II, DSIF, NELFからなる複合体の高次構造がX線結晶構造解析によって解明され,我々の生化学的な研究成果が見事に実証された15).DSIFとNELFによる伸長停止が遺伝子発現の重要な役割を果たすことが明らかになり,Pol IIの伸長停止・再開に関する我々の研究成果がやっと生命科学史上に残ることになった.
3)転写伸長停止・再開反応はPol II転写の律速段階である
伸長停止・再開は,当初は熱ショックやホルモンやサイトカインなどの外来刺激に応答性の遺伝子や病原性ウイルスなどの発現に関わる制御だと考えられたが12, 14, 16–19),MITのRick Youngらを含むいくつかの研究グループによるゲノムワイドな解析により,ほとんどすべてのPol II遺伝子の転写律速段階であることが証明された(図4B)20–23).したがって,高等真核生物における転写量の調節は,転写開始と伸長停止・再開の二つの段階で制御され,それにより時間および部位特異的に転写量が制御されている.ちなみに,NELFを発現抑制(ノックダウン)してPol IIの伸長停止が起こらないようにすると,外部刺激応答性の遺伝子は転写開始頻度が亢進されるが,伸長停止が起こらないので,転写開始頻度だけで転写量が決定される.すると,伸長停止・再開が起こる通常の転写と比べて,はるかに多量の転写産物の亢進が確認される.この意味するところは,外部刺激を介する開始段階での転写活性が非常に亢進して過剰量になっても,伸長段階の一時的な伸長停止からの再開頻度によって最終的に適度な転写量にうまく調節されることと,逆に,転写開始レベルで活性化されなくても,伸長停止・再開レベルで活性化が起こることにより転写量がうまく調節されることである.このような高等真核生物でしか起こらない転写の開始と伸長段階における二重の制御機構によって,緻密で高次な転写調節が行われていると考えられる(図4B).
今日,この伸長停止から再開する頻度を制御する因子をターゲットとした創薬開発が盛んに行われており,特定遺伝子の伸長停止・再開頻度を制御する新たな薬剤(後述のPROTACs/degraders)が脚光を浴びている.
このDRB作用機構の研究がきっかけとなり,ケミカルを用いて複雑な生体反応の制御機構を理解することに興味を持ち,ケミカルバイオロジー分野に足を踏み入れることになった.以下に,ケミカルバイオロジーに関する成果を紹介する.
5. サリドマイド催奇形性のターゲットの発見から新規薬剤開発への展開
1)サリドマイド催奇形性のターゲットの発見
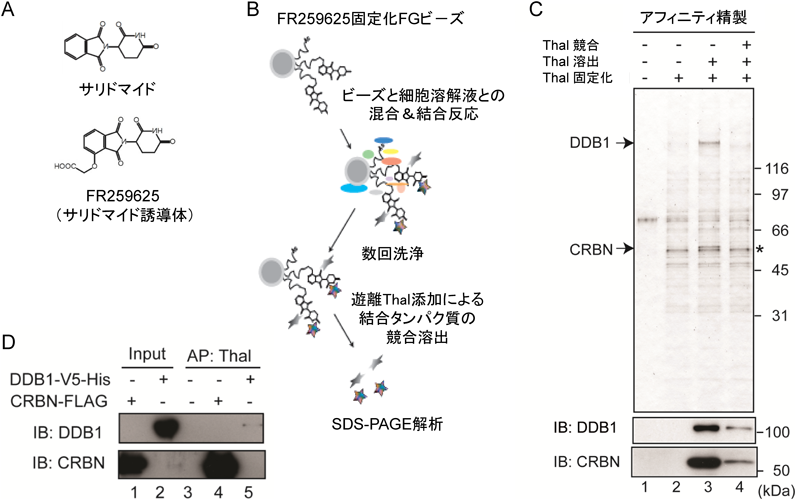
我々はこれまで数多くの化合物のターゲットを同定し,ターゲットが関わる複雑な生体反応の制御機構を解き明かしてきた.その中で最も輝かしい業績は,数十年以上も謎であったサリドマイド催奇形性のターゲットとしてセレブロン(cereblon:CRBN)を発見したことである28).この仕事は現東京医科大学准教授の伊藤拓水博士が主に行った.成果は2010年にScience誌のarticleとして掲載され,世界中のマスメディアで取り上げられ,一斉に報道され,国際的に大きな反響を呼んだ(図5).
サリドマイド(図5A)は1956年に有効な鎮静催眠剤として販売されたが,妊婦が服用すると生まれてくる子供にアザラシ肢症などの催奇形性が発症するという惨事を引き起こし,1962年には市場から完全に撤退した.ところが,30数年を経て,難治病であるハンセン病や多発性骨髄腫への有効性が認められ,現在では,厳しい統制の下,再び市場に舞い戻ってきたきわめて稀有な薬剤である29).
我々は,サリドマイドが持ついくつかの異なる薬理作用,たとえば,催奇形性,鎮静催眠作用,免疫調節作用,抗がん作用,血管新生阻害作用などに興味を持ち,2003年ごろから研究を開始した.その当時は,それら作用メカニズムはまったくわかってなかった.薬剤の作用メカニズムを理解するには,薬剤が体内で特異的に結合し,特定の薬理作用の要因となるターゲットを単離し,同定することが最善の策と考えた.そこで,サリドマイドの多様な作用メカニズムの理解を目的とし,独自開発のアフィニティビーズ技術を用いてサリドマイドのターゲットの単離・精製を試みた.
まず,サリドマイド誘導体をFGビーズ表面に固定化し(図5A),そのビーズと細胞破砕液とを混合して結合反応に数時間をかけ,よく洗浄した後に,ビーズ上の固定化サリドマイドと特異的に結合する二つのタンパク質を単離することに成功した(図5B).また,サリドマイドとの特異的結合を競合阻害実験や薬剤競合溶出実験を用いて証明することができた.次に,質量分析により,それらがCRBNおよびDDB1(damage-specific DNA binding protein 1)であると同定できた(図5C).さらに,それらの組換えタンパク質とサリドマイドとの結合をアフィニティビーズ技術で解析した結果,CRBNがサリドマイドと直接的に結合するが,DDB1はCRBNと会合して複合体を形成しており,サリドマイドとはCRBNを介して間接的に結合することがわかった(図5D).
また,生化学的手法を用いて,CRBNは直接会合するDDB1に加えて,Cul4やRoc1とともにタンパク質分解酵素であるE3ユビキチンリガーゼ複合体を形成し,ユビキチン化(Ub化)される基質タンパク質と選択的に結合する「基質受容体」であることを明らかにした.その後,サリドマイドはCRBNのC末端領域と結合することを示し,その領域の点突然変異体を作製した後に,サリドマイドとは結合しないが酵素活性を保持するサリドマイド非結合型CRBN変異体(YW/AA)を作製することに成功した.実験動物であるゼブラフィッシュおよびニワトリを用いて,この変異体がサリドマイド催奇形性を阻止することを明らかにし,CRBNがサリドマイド催奇形性の真のターゲットであることを証明することができた.加えて,サリマイドは,その代謝産物ではなく,そのものがCRBNと結合し,催奇形性発症に関わることを明らかにした.また,血管新生阻害がサリドマイド催奇形性の原因であるという仮説を否定することができた.さらに,CRBN E3ユビキチンリガーゼ複合体が四肢や耳の初期発達に関与し,サリドマイドはCRBNと結合し,その複合体の活性を変換することでそれらの初期発達異常を発症することを明らかにした28).
2)抗がん活性を持つ第二世代サリドマイド
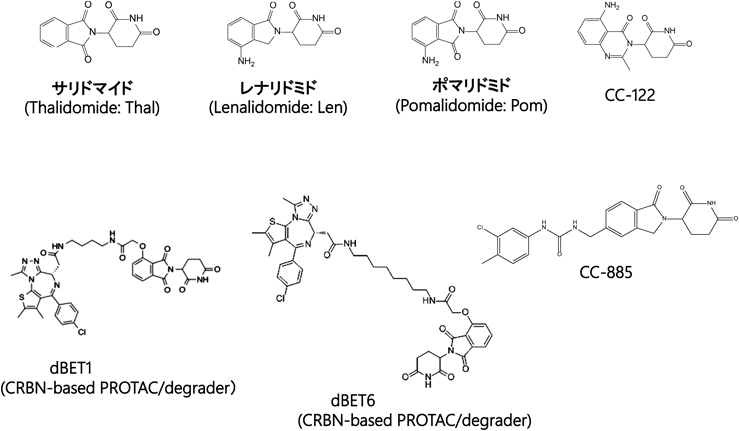
Science誌へのCRBNの論文掲載がきっかけとなり,米国製薬企業Celgene社との国際産学連携研究が始まった.Celgene社はサリドマイドを抗がん剤として薬剤市場に復帰させ,また,より優れた抗がん活性を持つ第二世代サリドマイドとしてレナリドミドやポマリドミドを開発し,販売していた(図6).それら3薬剤は,共通してT細胞の活性化を誘導する免疫調節作用を持つことからimmunomodulatory drugs,略してIMiDs,と呼ばれている30).IMiDsは,多発性骨髄腫(MM)に対して多面的な治療効果を発揮する優れた抗がん剤である.がん細胞に対しては細胞増殖を阻害する抗がん効果を示し,さらに免疫担当細胞に対してはIL-2の発現誘導を介して細胞障害性T細胞(CTL)を含むT細胞を活性化して,抗がん効果を増強できる.Celgene社は,我々が発見したCRBNに大変興味を持ち,論文発表してからあまり時間が経たないうちに連絡がきて,西海岸のサンディエゴにある新たな研究所に招聘され,IMiDsの抗がん効果におけるCRBNの役割に関する共同研究が始まることになった.
3)IMiDsの抗がん作用におけるCRBNの役割
結論から先にいうと,Celgene社との共同研究によって,CRBNがサリドマイドの副作用だけでなく,IMiDsの主作用である抗がん作用のターゲットでもあることがわかった31, 32).
我々はまず,MM由来の細胞株を用いて,CRBNを発現抑制(ノックダウン)したときにIMiDsの感受性が変わるか否かを試みた.通常のMM細胞株ではIMiDs存在下では細胞増殖が阻害され,IMiDs感受性を示すが,CRBNを発現抑制するとIMiDsに対して耐性になる.これはまさにCRBNがIMiDsによるがん細胞の増殖阻害に関わっていることを示すものである.次に,IMiDs存在下でMM細胞の長期培養を続けると,IMiDs存在下でも増殖し続けるIMiDs耐性のMM細胞が出現する.その耐性細胞ではCRBNの発現が著しく低下しているので,MM細胞がIMiDs感受性を示すにはCRBNの存在が必須であることがわかった.
さらに,IMiDsによるMM細胞の増殖阻害を分子レベルで解析した.MM細胞の増殖はIRF-4とc-mycに依存的で,MM細胞をIMiDs処理すると,その増殖・生存に必須なc-myc遺伝子やIRF-4遺伝子の発現が抑制されることがすでに報告されている33).そこで,CRBNを発現抑制したMM細胞を用いて,IMiDsによるc-mycやIRF-4遺伝子の発現抑制を検討した.その結果,CRBNを発現抑制するとIMiDs処理によるc-mycおよびIRF-4の遺伝子発現は抑制されず,がん細胞増殖も阻害されないことから,IMiDsによるMM細胞の増殖阻害作用はCRBN依存的であることがわかった32).興味あることに,IRF-4とc-mycの遺伝子発現は,相互に調節し合っており,片方の発現が低下すると他方も低下し,逆に片方が増えると他方も増える仕組みになっている.
加えて,ヒトCRBNとIMiDs複合体の高次構造をX線結晶構造解析によって明らかにした.IMiDsに共通した部分構造であるグルタルイミドがCRBNのC末端領域にある三つのトリプトファンで形成されるポケット(Tri-trp pocket)に入り込むことがわかった34).この結果は,他グループによるニワトリCRBNとIMiDs複合体の高次構造の解析結果と同じで,互いの確実性が増強された35).
4)光学異性体を持つIMiDs
サリマイドを含むすべてのIMiDsは,S体とR体と呼ばれる光学異性体を有している.これまでサリドマイドのS体とR体の機能を識別する科学的な方法がなく,ほとんどの人がS体が副作用,R体が主作用に関与すると誤った思い込みをしていた.我々は,サリドマイドのターゲットであるCRBNを用いて,S体とR体の機能を科学的に区別することが可能になったので,長年の誤解を払拭し,真実を明らかにすることができた.
まず,S体またはR体IMiDsとCRBNとの相互作用を生化学的に解析した.その結果,すべてのIMiDsのS体は,R体と比べてはるかに強く,しかも安定的にCRBNと結合することがわかった.その結果は,CRBNとIMiDsからなる複合体のX線結晶構造解析によっても証明された34, 36).さらに,副作用である催奇形性発症および主作用である抗がん効果に関しても,R体と比較するとS体の方がはるかに高いことを明らかにした.したがって,S体IMiDsが主作用にも副作用にも関与することがわかった.
また,IMiDsの光学異性体は,生理的条件下ではS体からR体へ,R体からS体へと異性化(ラセミ化)し,ラセミ体と呼ばれるS体とR体が等量存在する状態になることがすでに報告されている.ちなみに,ゼブラフィッシュは弱酸性条件下(~pH 6.0)で飼育でき,弱酸性条件下ではラセミ化が起こりにくく,光学異性体による催奇形性実験にゼブラフィッシュは適している.
したがって,ヒト体内でラセミ体であるIMiDsがCRBNと出会った場合には,S体が優先的にCRBNと結合し,残るR体は異性化してラセミ体となり,S体が常に補充されるので,光学異性体はIMiDsの薬効にはさほど影響しないことが明らかになった.
5)IMiDsによる抗がん作用のメカニズム
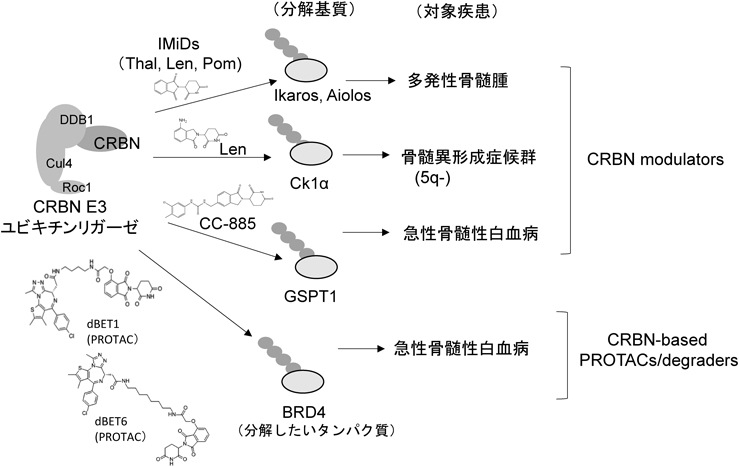
IMiDsの抗がん作用メカニズムを理解するために,IMiDs存在下でCRBNと結合し,Ub化が亢進される基質タンパク質をスクリーニングした.MM細胞をIMiDsで処理し,適当な処理時間後に細胞を破砕し,破砕液中の総タンパク質をプロテアーゼで分解し,Ub化されたペプチドを特異抗体を用いて濃縮・回収した後に,タンデム質量分析によって,IMiDs処理でUb化が亢進したペプチドを網羅的に同定した.最も亢進したものとして血球系転写因子であるAiolosを同定することができた.Aiolosは,Ikarosファミリーに属する転写因子で,そのファミリーには五つのメンバーが存在し,その中の三つ,すなわち,Ikaros, Aiolos, Heliosが血球系細胞で特異的に発現している.そこで,それらメンバーがIMiDs存在下でCRBNの新規基質になりうることを確かめるために,健常人から調製したT細胞画分をIMiDsで処理した.すると,IkarosとAiolosはCRBN依存的にUb化され,プロテアソームにより分解されるが,HeliosはUb化も分解もされないことがわかった.CRBN依存的とは,CRBNを発現抑制するとUb化・分解されないことを意味する.
また,IkarosとAiolosは,IMiDs非存在下ではCRBNと結合しないが,IMiDs存在下で,CRBNと選択的に結合することがわかった.この結果は,IkarosおよびAiolosは,IMiDsがCRBNと結合したIMiD・CRBN複合体を認識し,結合して,新規基質としてUb化され,プロテアソームで分解されることを示している37, 38).というのは,IMiDs非結合型CRBNYW/AA変異体は,IkarosやAiolosをUb化・分解できないからである.
また,IMiDs処理によりIkarosおよびAiolos(両者はIL-2の転写レプレッサーであることが既知である)がUb化・分解され,減少すると,免疫担当T細胞ではその抑制的支配下にあるIL-2遺伝子の発現は抑制解除され,亢進し,それによってT細胞活性化が誘導され,免疫調節作用が発現されることがわかった.一方,MM細胞ではIkarosおよびAiolos(両者はIRF-4の転写アクチベーターであることが公知である)が分解されると,その促進的制御下にあるがん細胞増殖・生存に必須なIRF-4やc-mycの発現が減少することで,がん細胞の増殖が抑制されることがわかった.これがIMiDsによる抗がん作用のメカニズムで,IMiDsは,がん細胞に対する増殖・生存阻害作用と,免疫担当細胞に対するT細胞活性化作用という多面的な治療効果を発揮する優れた抗がん剤である(図7)39).
また,IMiDsの中でレナリドミドだけは,骨髄異形成症候群(MDS)の5q-症候群に対する特効薬で,その興味ある作用メカニズムが最近解き明かされた40).
レナリドマイドがCRBNと結合して複合体を形成すると,新規基質としてカゼインキナーゼ1α(CK1α)がリクルートされ,Ub化・分解されることがわかった40).通常では,CK1αはがん抑制因子p53をリン酸化して,不活性化する.CK1αは5番染色体の長碗(5q)にコードされている.5q-症候群では二倍体染色体の片方の5q部位が欠損して一倍体となっているために,CK1αの発現は通常の半分量である.さらに,半分量に減ったCK1αは,レナリドミド処理によってUb化・分解され激減するので,p53のリン酸化・不活性化は起こらず,CK1αによりリン酸化されない活性型p53が蓄積・増加する.その結果,細胞アポトーシスが誘導され,治療効果が発現されるというメカニズムが明らかになった.ちなみに,通常の二倍体細胞では,レナリドミドによるCK1αのUb化・分解が起こっても,合成量が確保されるのでCK1αは激減することはない.それゆえ,活性型p53の蓄積は起こらず,がん細胞のアポトーシス誘導には至らない.以上が,レナミドミドが5q-症候群の特効薬である主な理由である(図7).
この研究を通じて,一つ重要な点が明らかになった.それは,レナリドミドの有効な治療効果を得るには,基質タンパク質の発現量,すなわち,タンパク質の合成と分解の割合を考慮すべきということである.これは,今後,タンパク質分解系の薬剤開発を目指す上で,大事な情報である.
さらに近年,レナリドミドは成人T細胞白血病/リンパ腫(ATL)の治療薬としてわが国で認可された.しかし,その治療効果のメカニズムはいまだわかってない.
これまでの成果によって,IMiDsがCRBNと結合してIMiDs・CRBN複合体を形成すると,CRBNの基質選択性が変化し,その複合体との結合に特異性および親和性が最も高い基質タンパク質がリクルートされ,CRBN E3ユビキチンリガーゼによってUb化・分解されることがIMiDsの薬理作用機序の本質であることがわかった(図7)41).これまでの研究により,下記の「CRBN modulators」と「CRBN-based PROTACs(タンパク質分解誘導薬)」と呼ばれる二つのタイプの新規抗がん剤開発への道筋が切り開かれた(図7)42, 43).
6)CRBN modulatorsへの発展
近年,Celgene社により第三世代サリドマイドとしてCC-122やCC-885などが開発された(図6).CC-122はびまん性大細胞型B細胞リンパ腫(DLBCL)に有効な抗がん剤だが,いまだに基質タンパク質は同定されていない44).一方,CC-885は細胞毒性が強く,当初は薬剤には不向きだとも思われたが,我々はCC-885がCRBNと結合すると,新規基質として翻訳終結制御因子GSPT1がCRBN上にリクルートされ,Ub化・分解されることを見いだした42).これにより,CC-885は再検討され,きわめて低濃度(nM以下)で急性骨髄性白血病(AML)に対して有効な治療効果を発揮することがわかり,CC-885とその誘導体がAML治療薬として開発されている.我々は,CC-885の治療効果のメカニズムを解き明かすとともに,CC-885がCRBNと基質GSPT1との間で,これまでになく強固な複合体を形成することを見いだした.これならば,X線結晶構造解析によって複合体の高次構造が解析できると直感し,すぐに構造解析を始め,DDB1/CRBN/CC885/GSPT1の四者からなる複合体の高次構造をX線結晶構造解析および電子顕微鏡解析によって解明することができた42).
第三世代サリドマイドは,第二世代サリドマイドからなるIMiDsの範疇には収まらない.そこで,CRBNと結合して固有の新規基質をCRBN上にリクルートし,E3ユビキチンリガーゼ活性によりUb化・分解することで治療効果を発揮するサリドマイドおよびその合成展開した誘導体(第二世代や第三世代のサリドマイドを含む)のすべてを「CRBN modulators」と総称することになった(図7)29).現在,CC-122やCC-885以外の有効なCRBN modulatorsがいくつかすでに開発されている45).
7)CRBN-based PROTACs/degraders(タンパク質分解誘導薬)への展開
2001年にエール大学のCraig Crews教授が,二つの機能を有する小分子で,ターゲットタンパク質とユビキチンリガーゼの両方に結合して,ターゲットタンパク質をユビキチンリガーゼの近傍に引き寄せて基質としてUb化・分解するキメラ薬をターゲットタンパク質分解誘導薬(proteolysis targeting chimeras:PROTACs)と名づけた.CRBNはまだ発見されておらず,VHLユビキチンリガーゼを利用したPROTACsが,これまで研究開発されてきた46).
その歴史的背景に基づき,ハーバード大学の研究グループがCRBN E3ユビキチンリガーゼを利用したCRBN-based PROTACsを作製し,効果を検討してみると,優れた分解活性が得られた.CRBNと結合するサリドマイドに,興味あるタンパク質(protein of interest:POI)と結合する薬剤を連結したキメラ薬を合成し,POIをCRBN近傍にリクルートし,Ub化・分解するCRBN-based degraders(彼らはPROTACsではなくdegradersと呼ぶ)の研究開発が本格化し,現在,盛んに行われている.
このPROTACs/degradersの開発によって,化合物をデザインすることで任意のタンパク質をUb化・分解することが可能となった.これによって,従来は創薬の開発につながらないようなundrugabbleなタンパク質と考えられていた転写伸長制御因子などをターゲットとすることが可能になった.
そこで,上記の我々の基礎研究から見いだされたとおり,転写伸長停止・再開段階が転写律速段階であることがわかったので,世界をリードする米国ボストンの研究グループが大きな製薬企業と共同して転写伸長停止・再開の制御因子を創薬ターゲットとしたdegradersの開発を積極的に行っている.たとえば,がん遺伝子であるc-myc遺伝子の転写伸長停止・再開反応の制御因子であるBRD(bromo-domain-containing protein)4の阻害剤JQ1が開発されたが,最近JQ1よりも,より高い薬効を発揮するJQ1とサリドマイドを連絡したキメラ剤dBET147)や,それをさらに改良したdBET6が開発された(図6)48).dBET6はBRD4に加えて,BRD2やBRD3も分解し,c-myc遺伝子発現をより厳しく抑制することで抗がん活性(急性骨髄性白血病の治療など)を示すことから,dBET6はc-myc遺伝子増幅を認めるがん種などへの抗がん剤として臨床応用が期待されている.また,近年,転写伸長停止から再開を誘導するP-TEFbのキナーゼサブユニットであるCDK9をターゲットとしてUb化・分解するCRBN-based degraderが開発された.それら開発された伸長停止・再開反応の制御因子をターゲットとしたdegradersは,がんの治療薬として開発されているが,転写伸長停止・再開反応の制御機構をより詳細に解明している基礎研究においても大いに役立っている(図7).
また,転写伸長停止・再開の制御因子に加えて,多様ながん関連因子,たとえばBCR-ABLなどをターゲットとしたCRBN-based degradersが世界中で開発され始めている.