ロドプシンの作動メカニズムMechanism of Rhodopsin Function
名古屋工業大学大学院工学研究科生命・応用化学科Nagoya Institute of Technology ◇ 〒466–8555 名古屋市昭和区御器所町 ◇ Showa-ku, Nagoya 466–8555, Japan
発行日:2019年8月25日Published: August 25, 2019
ロドプシンはレチナール分子を光吸収に用いて光情報変換やエネルギー変換に働く膜タンパク質である.動物ロドプシン(animal rhodopsin)は視物質としてGタンパク質を活性化する一方,微生物ロドプシン(microbial rhodopsin)はポンプ・チャネル・センサー・酵素など多彩な機能を持つ.ロドプシンは構造機能相関の研究が進んだ膜タンパク質としてよく知られているが,最近では光遺伝学(optogenetics)の主要ツールとして応用研究への期待も高い.我々はこれまでロドプシンの作動メカニズムを研究する中で,新しいロドプシン機能の発見や創成にも関わってきた.本稿では,ナトリウムポンプや内向きプロトンポンプ,ヘリオロドプシンなど我々の微生物ロドプシン研究を中心にロドプシンの作動メカニズムについて最新の知見を紹介する.
© 2019 公益社団法人日本生化学会© 2019 The Japanese Biochemical Society
植物の光合成,動物の視覚に代表されるように,光は我々の生存に必須であり,「いのちの源」として大きな恵みを与えてくれる.光受容タンパク質は,生物が光を利用する際に受容体となるタンパク質であり,光を吸収するための低分子(chromophoreあるいは発色団)がタンパク質という限定された環境のもとに存在する.そして光を吸収すると多くの場合,タンパク質場に特異な光化学反応(異性化,電子移動など)が起こる結果として機能が発現する1, 2).
光受容タンパク質の中で,本稿で紹介するロドプシンは,動物が持つ動物ロドプシン(animal rhodopsin)と細菌などの微生物が持つ微生物ロドプシン(microbial rhodopsin)の二つに分類することができる3).いずれもαへリックスが膜を7回貫通した構造を持つ膜タンパク質である(図1).両者にアミノ酸の相同性はなく,別々の進化をたどってきたと考えられているが,動物ロドプシンと微生物ロドプシンには共通点も多い.たとえば,ロドプシンの発色団であるレチナール分子は7番目のへリックスにあるリシン残基とシッフ塩基結合を形成する(図1).ニンジンなどに含まれるβカロテンを半分に切ったのがビタミンAであるが,レチナールはそのアルデヒド型であり,種々の異性体構造の中で最も安定なものがall-trans型レチナールである(図2)3).微生物ロドプシンはall-trans型レチナールを発色団とするが,動物ロドプシンは11-cis型レチナールを発色団とする.11-cis型レチナールは熱安定性が低く,溶液中では容易に熱異性化してall-trans型になってしまうが,動物は網膜中にall-trans型を11-cis型へと転換する異性化酵素を持ち,動物ロドプシンは11-cis型レチナールを安定に結合するポケットを持っている4).
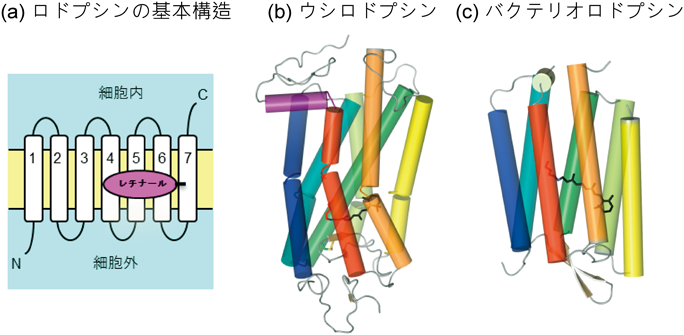
いずれもαヘリックスが膜を7回貫通した構造を持っており,C末端側が細胞質に面している.文献3の図を改変.
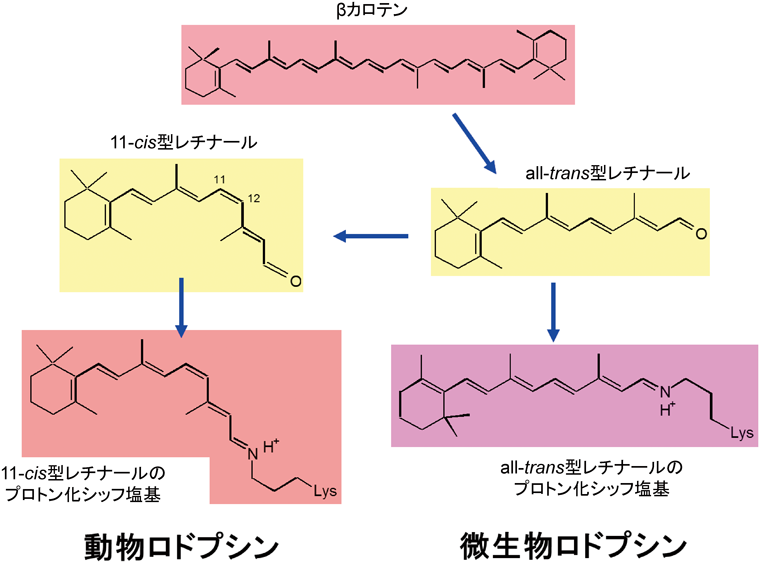
動物ロドプシンは11-cis型レチナール,微生物ロドプシンはall-trans型レチナールが第七ヘリックスのリシンとシッフ塩基結合を形成する.レチナールシッフ塩基は紫外線領域に吸収を持つが,シッフ塩基がプロトン化し正電荷を持つことで可視領域の吸収が実現する.
動物ロドプシンはいずれもGタンパク質共役型受容体(GPCR)としてホルモンや神経伝達物質の受容体の仲間である.ヒトの場合,明暗視に働くロドプシン(狭義の「ロドプシン」)や3種類の色覚視物質,さらにはこれら視覚の光センサーと相同性を持ったメラノプシンなど数種類の動物ロドプシンを持っている.動物ロドプシンが水溶性の三量体Gタンパク質を活性化することによる光情報伝達という唯一の活性化機構を持つのに対して,微生物ロドプシンは後述するように実にさまざまな働きを持つ.私は京大大学院理学研究科・生物物理の吉澤透教授の研究室で動物ロドプシンと出会い,視覚の初期過程がレチナールの異性化反応であることを示して学位を取得した5, 6).以来,ロドプシンが示す反応ダイナミクスの美しさに魅了され,少しでもその働きを理解しようと努力を続けている.本稿では,我々の微生物ロドプシン研究に焦点を絞りながら,ロドプシンの作動メカニズムについて紹介したい.
目の光センサーとして古くからその存在が知られていた動物ロドプシンと違って,微生物ロドプシンの歴史はさほど古くない.微生物ロドプシンの中で最初に発見されたのが有名なバクテリオロドプシン(BR)であり,塩濃度が飽和するような極限環境で成育できる高度好塩菌の紫膜から1971年に発見された7).BRは光駆動外向きプロトンポンプとして機能し,生成されたプロトンの濃度勾配がATP合成のために利用される.高度好塩菌という特殊な細菌に存在するBRであるが,膜タンパク質研究の歴史においては特別な位置を占めてきた.膜タンパク質の構造情報が皆無であった時代に,英国のHendersonらは電子線を用いた解析によりBRが膜中でヘリックス構造を形成していることを明らかにした8).αヘリックスが膜を貫通するという現在の常識が初めて明らかにされたのがBRであり,42年後の2017年にHenderson博士はノーベル化学賞を受賞する.一般に膜タンパク質を図示するときには細胞外側を上に描くが,ロドプシン分野の研究者は慣例として細胞外側を下に描いており,これはHenderson博士が最初に置いた向きに由来する9).最も理解の進んだ膜タンパク質の一つとして,光反応中間体の構造解析10)や最近では自由電子レーザーを用いた構造ダイナミクス解析11)など,新しい手法が最初に試みられるタンパク質である.
BRに代表される微生物ロドプシンが動物ロドプシンと異なる大きな特徴として,光反応サイクルをあげることができる.動物ロドプシンであるウシやヒトのロドプシン中でレチナールが光を吸収すると11-cis型からall-trans型に異性化反応が起こり,さまざまな中間体を経て最終的にall-trans型レチナールはタンパク質から解離する(図3).網膜中では新しい11-cis型レチナールが供給されるが,in vitroでは一度しか光を当てられないため暗室実験が必要となる.一方,BRなどの微生物ロドプシンが光を吸収すると,all-trans型から13-cis型に異性化反応が起こり,さまざまな中間体を経てレチナールはall-trans型へと熱異性化する(図3).動物ロドプシンと微生物ロドプシンのこのような光反応性の違いは,レチナールの異性化における構造変化の程度に由来するものと考えられる.
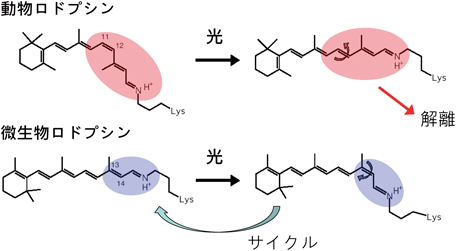
光を吸収すると,動物ロドプシンでは11-cis型レチナールのC11=C12の二重結合が,微生物ロドプシンではall-trans型レチナールのC13=C14の二重結合が異性化する.
BRはJ, K, L, M, N, Oと名づけられた色の異なる中間体を経由する10ミリ秒程度の光反応サイクルの間に,細胞質側から細胞外側に1個のプロトンが玉突き的に輸送される3).中性付近では4個の結合基(細胞質側から順に,D96,レチナールシッフ塩基,D85, E204を中心とした水クラスター)を経由する5回のプロトン移動によって一方向へのベクトル的輸送が実現するのである.光反応サイクル中で鍵となる中間体K, M, Nを図4に示した.対イオンX−はBRではD85であり,内部の水素結合構造は細胞外側領域が開いている(EC open).光吸収後,10−13秒の時間領域で異性化反応が起こり,13-cis型に異性化したK中間体を生成する.次に起こるのがプロトン移動反応であり,M中間体の生成時にシッフ塩基から対イオンに移動するが,超高速光異性化(10−13秒)に対してこの反応は10−5秒と,8桁も遅い時間を要してプロトン移動が起こる.ポンプの一方向性に関係して重要なことは,プロトンがD85にわたった後,反対側からプロトンを受け取ることであり,玉突き的なプロトン移動が起こる.実際に,シッフ塩基の再プロトン化は逆側(D96)から起こり(図5),このとき疎水的な細胞質側領域に輸送経路を構成するため,ヘリックスが外向きの開運動を起こす(CP open).最後にレチナールが13-cis型からall-tran型に熱異性化して光反応サイクルが完結し,1回の光反応サイクルで1個のプロトンを細胞外にポンプする.図4は,異性化とプロトン移動という異なる反応を組み合わせることで一方向のプロトン輸送という機能が発現することを示している.機能発現において重要なことは時間的・空間的な階層性を実現することであるが,四つのステップはそれぞれ10−13秒,10−5秒,10−3~10−2秒,10−2~10−1秒と分離された時間領域で起こり,ベクトル性を保証している.
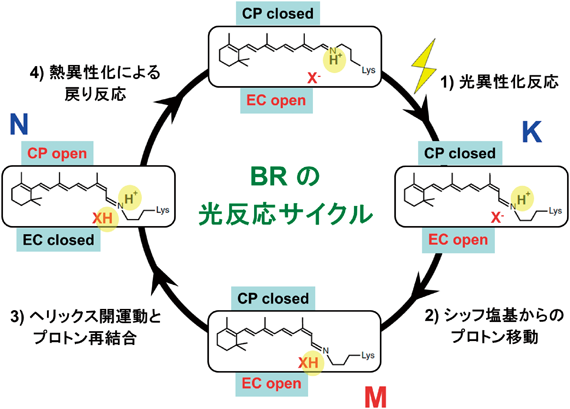
X−はレチナールシッフ塩基の対イオンであるD85を示す.CP, ECは細胞質側,細胞外側を表し,水素結合ネットワークによりシッフ塩基部位とつながっている場合,openと表記している.文献13の図を改変.
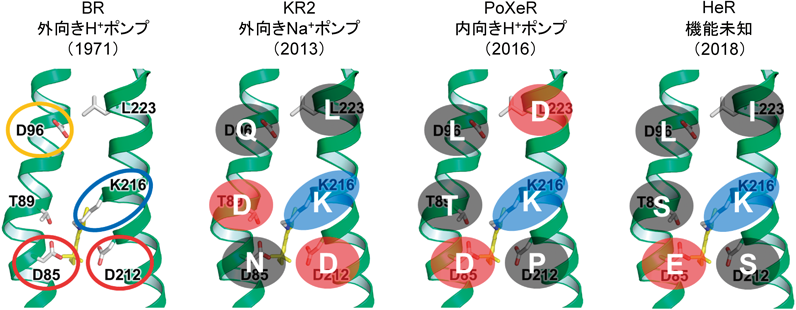
左から外向きプロトンポンプのバクテリオロドプシン(BR),外向きナトリウムポンプ(KR2),内向きプロトンポンプ(PoXeR),ヘリオロドプシン(HeR)において重要なアミノ酸残基を,BRの構造上に示した.上が細胞質側,下が細胞外側であり,それぞれ左がCヘリックス(TM3),右がGヘリックス(TM7)である.BRのK216にレチナールが結合するが,リシンとの結合はすべてのロドプシンに共通である(正電荷を青色で示す).CヘリックスのD85はプロトンアクセプター,D96はプロトンドナーとして働き,T89と合わせてDTDモチーフと呼ばれる.BRのD85, D212は負電荷を持つ一方(赤丸),D96はプロトン化している(黄丸).ナトリウムポンプはNDQモチーフを持ち,BRからアスパラギン酸が細胞質側にヘリックス1ターン移動した形になる.内向きプロトンポンプはBRにおけるD212の位置が中性化される一方,L223位のアスパラギン酸がプロトンアクセプターとして働く.D212位のアスパラギン酸は微生物ロドプシンで高度に保存されているが,HeRも中性残基(セリン)であり,図の上部は疎水的な残基で構成される.
私は京大大学院理学研究科・生物物理の前田章夫教授の研究室で助手になったとき,BRに結合した水分子の赤外分光解析を開始した.京大時代の仕事をまとめた総説12)はレニンジャーの『生化学』で紹介されており,水分子を含む水素結合ネットワークの解析における赤外分光法の重要性を示している.2001年に名工大で自分の研究室を持ってからはさらに計測の精度を高め,すべての水素結合強度の水分子を捉えることに成功することができた13).これらの仕事は助教の古谷祐詞博士(現名工大准教授),ポスドクのV.A Lorenz-Fonfria博士(現バレンシア大准教授),大学院生であった柴田幹大博士(現金沢大准教授)らとともに行った.
1971年のBRに続き,高度好塩菌からは1977年に内向きクロライド(塩化物イオン)ポンプであるハロドプシン(HR)14),1982年に正の走光性センサーであるセンサリーロドプシンI15),1985年に負の走光性センサーであるセンサリーロドプシンII(フォボロドプシン)16)が発見され,高度好塩菌は4種類の異なる機能を持ったロドプシンを保有することが明らかとなった.ポンプという機能に加えて,2種類のセンサリーロドプシンは膜結合型の伝達タンパク質を活性化することで情報伝達を行うことが明らかになり,動物ロドプシンとは異なった光信号変換機構が存在することがわかった3).興味深いことに,センサリーロドプシンは膜結合型の伝達タンパク質が存在しないとプロトンポンプとして機能することが知られており17),プロトンポンプ機能がこれらのタンパク質の起源であったと推測されている.この中でドイツ人によって発見されたBR以外の3種類の発見においては,日本人研究者の貢献が多大であったことを特記しておきたい.日本が世界をリードしてきた動物ロドプシンの研究と合わせ,このような歴史と伝統が日本のロドプシン研究には息づいている.
生物は真核生物,古細菌,真正細菌の3種類に分類されるが,BRなどを有する高度好塩菌は古細菌に属するため,これらのタンパク質群は当初,古細菌ロドプシン(Archaeal rhodopsins)と呼ばれていた.面白いタンパク質であることは認められていたものの,塩辛い湖に生息する古細菌のみが持つ膜タンパク質であるため脚光を浴びることはなかった.この状況を一変させたのはゲノム科学の進展である.古細菌だけでなく,カビ類や藻類などの真核生物や真正細菌から次々にBRのようなタンパク質が発見され,現在では微生物ロドプシンと総称されるようになったのである.この中でも圧倒的な割合を占めるのが海洋性細菌から発見された一群であり,今や海にいる細菌の7割以上はロドプシンを持つことが示唆されている.海水を手ですくったとき,その中に含まれる生物の7割以上がロドプシンを持つという事実は,発見当初のBRの位置づけと比較すると感慨深いものがある.これらは最初に発見された細菌の名前をとってプロテオロドプシン(PR)と総称されるが18),そのほとんどがBRのように光駆動プロトンポンプとして働く.つまりPRはBRのD85, D96(図5)と同様に,負電荷を持ったプロトンアクセプターとプロトン化したドナー(PRではグルタミン酸)を持つ.PRを持つ細菌は食物連鎖の最下位に位置し,分解者として従属的な栄養摂取のみを行っているとこれまで考えられてきたが,光駆動プロトンポンプを用いて太陽の光を積極的に利用していることが判明したのである.
PRの発見は2000年であるが,2002年,緑藻のクラミドモナスから発見されたチャネルロドプシンは,それまでに知られていたBRやPRのようなポンプではなく,濃度勾配に従って陽イオンを輸送するチャネルであった19).それから5年も経たないうちに,米国のDeisserothら20)や日本の八尾ら21)が,チャネルロドプシンを特定の神経細胞に発現させて,光で神経興奮を制御するという技術を開発した.この手法は,Deisserothらにより光学(Optics)と遺伝学(Genetics)を組み合わせてオプトジェネティクス(Optogenetics;光遺伝学)と呼ばれるようになった22).オプトジェネティクスにおいては,クラミドモナスのチャネルロドプシン2(ChR2)が光を吸収するとチャネルが開きナトリウムイオンが流入し,その結果,神経細胞が脱分極することで活動電位を発生することになる(図6).脳神経生理学者がそれまで行ってきた神経細胞の電気刺激では特定の細胞だけを刺激するのが困難であったため,オプトジェネティクスは爆発的な広がりをもたらすことになった.この技術が開発された当時,ChRのようなチャネルと違って1個の光子で1個のイオンしか輸送できない光駆動イオンポンプは応用が困難であると私は考えていたが,クロライドポンプであるHR23)やBRなどのプロトンポンプ(BRと高い類似性を持つ古細菌型ロドプシンがよく利用される)24)も細胞を過分極させることで神経抑制に応用可能であることが明らかとなった.イオン輸送性ロドプシンはオプトジェネティクスのツールとして期待が高く,図6に示すとおり,後述する新しいロドプシンもすぐにオプトジェネティクス応用が試みられている.
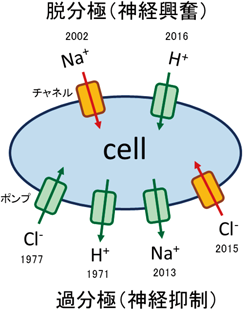
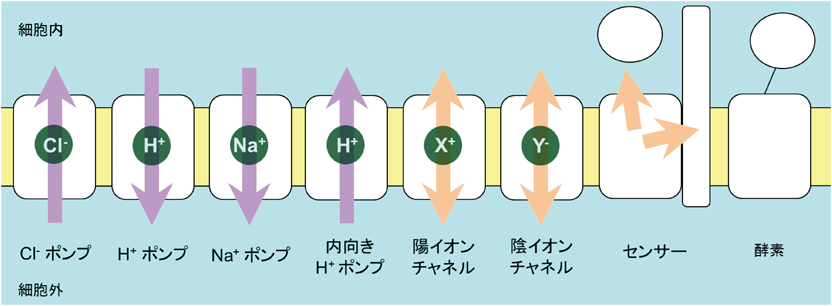
これらのロドプシンに光を照射すると,陽イオンチャネルと内向きプロトンポンプは脱分極,内向き陰イオンポンプと外向き陽イオンポンプ,陰イオンチャネルは過分極をもたらす.これらのロドプシンが発見された年を,輸送されるイオンとともに示す.
ここであらためて,動物の行動を制御するオプトジェネティクスに,動物ロドプシンではなく微生物ロドプシンが使われた理由を整理しておこう.まず,動物ロドプシンの11-cis型レチナールと異なり,発色団であるall-trans型レチナール(図2)が脳などに十分量,存在することがあげられる.このため,発色団分子を外から加える必要がない.第二の理由として,動物ロドプシンと異なり,光反応がサイクルを示す(図3, 4)ことがあげられる.さらに第三の理由として,光で能動的あるいは受動的にイオンを輸送するため,神経細胞の脱分極や過分極をもたらすことができる.このような性質ゆえ,微生物ロドプシン遺伝子を標的とする細胞に発現さえすれば,光による繰り返しの細胞応答が実現したのである.ロドプシンのメカニズム解明という基礎研究に突き進んできた我々にとって,オプトジェネティクスの利用者からツール開発が期待される状況は興味深いものがある25, 26).我々はこれまで赤外分光を中心にロドプシンの分光研究を進めてきたが,電気生理学的手法は必須であり,ドイツから加わった角田聡博士(現名工大特任准教授)を中心に研究を行っている.
ナトリウムイオンとプロトンは生命活動に中心的に働く陽イオンであるが,光駆動ポンプはプロトンだけであり,ナトリウムポンプは存在しないというのが常識であった.光吸収を担う発色団が正電荷を持つためナトリウムイオンなどはレチナール近傍に結合できず,結合できなければ光のエネルギーを使って輸送できないと考えられてきたのである.実際にプロトンポンプやクロライドポンプの基質は,光を吸収する前にレチナールシッフ塩基に共有結合している(プロトン)か,対イオン(クロライドイオン)として結合している.私は講演などでナトリウムポンプが存在しない理由を活性中心であるレチナールシッフ塩基部位の静電相互作用により明快に説明していたのであるが,BRの発見から40年以上経過してから光駆動ナトリウムイオンポンプの存在がわかった27).この発見をもたらしたのは,第三ヘリックスを構成するアミノ酸配列の特異性である.BRのプロトンアクセプターであるD85はシッフ塩基の正電荷を安定化させる対イオンとして働くが,ヘリックス1ターンだけずれた位置(BRのT89位)にアスパラギン酸を持つロドプシンが海洋性細菌Krokinobacter eikastusに見いだされ,その性質を調べた結果,新たな機能に行きあたったのである(図5).第三ヘリックスのD85, T89, D96に対応する残基はモチーフと呼ばれ,DTDまたはDTEであればプロトンポンプ,NDQであればナトリウムポンプ,カルボン酸を持たなければ(NTQまたはTSA)クロライドポンプであることがわかっている28).
ではどうやって光駆動ナトリウムポンプは機能するのだろうか? 陽イオンを変えても色が変わらなかったことから予想どおり,正電荷を持ったレチナールの近傍にナトリウムイオンは結合しないものの,我々はまず赤外分光を用いてナトリウムイオンの結合部位が出口側部位に存在することを見いだした(図7)27).ところが驚いたことに結合部位を変異で欠損させてもナトリウムポンプ機能は保たれていた.輸送にエネルギーを必要とするポンプはイオンを内部に結合している状態でエネルギー入力と共役して輸送する例がほとんどであり,パナマ運河のような二つのゲートによる船の出入りとエネルギー入力(運河の水位上昇)によって説明できる29).しかしながら,プロトンやクロライドイオンを輸送するロドプシンと異なり,ナトリウムイオンを結合していなくてもポンプできるのだ.この事実から我々は,ナトリウムイオンを細胞内から取り込む過程は拡散により支配される受動的なものと解釈している.ナトリウムイオンが取り込まれるとレチナールシッフ塩基近傍に過渡的に結合するが,ナトリウムイオンとプロトンが競合するモデルで取り込みがよく説明できることは受動的な輸送過程を支持している30).プロトンポンプから進化の中で競合してナトリウムイオンをポンプする装置を作り,それが400 mM Na+,pH 8という海水中で7桁多いナトリウムイオンをポンプするようになった可能性がある.
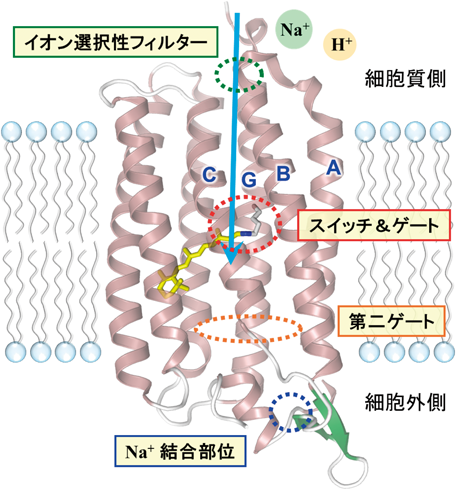
機能に重要な部位を構造上に示した.文献29の図を改変.
光駆動ナトリウムイオンポンプはリチウムイオンも一方向に輸送できるが,カリウムイオンやセシウムイオン中ではプロトンを輸送する27).既述のとおり,細胞内にはカリウムイオンが多いため,ナトリウムイオンだけをポンプしてカリウムイオンの濃度勾配を変えないという事実は理にかなっている.一方,ナトリウムイオンの方がカリウムイオンよりも水和力が強いため,水和イオンサイズはナトリウムイオンの方が大きいことに注意が必要である.この事実は,ナトリウムイオンの一方向輸送において必ず脱水和が起こることを示唆している.ではタンパク質内部で水分子を引きはがし,直径1.9 Åのナトリウムイオンを通して2.8 Åのカリウムイオンを通さない選別部位はどこにあるのだろうか? 我々は東大・濡木研と共同でX線結晶構造解析を行ったところ,イオン取り込み部位に狭くなっている部分を見いだしたため,この部位に変異を導入した結果,カリウムイオンをポンプするタンパク質を創成することに成功した31).我々はさらに入口部位の変異を試すことで,ルビジウムイオンとセシウムイオンの一方向輸送を達成することができた32).このタンパク質は世界で初めてセシウムイオンの一方向輸送を実現したものであり,福島原発事故で放出された137Cs+の回収につながる可能性を秘めた基盤技術である.
光駆動ナトリウムポンプに関する現在の理解を図7に示した.出口側表面に存在するナトリウムイオン結合部位は能動輸送には必須ではなく,タンパク質の安定性に関わる.入口側表面に存在するイオン選択性フィルターにより大きなイオンが排除される一方,ナトリウムイオンはプロトンと競合して取り込まれる.解析の結果,プロトン取り込み速度がナトリウムイオンより8000倍速いことがわかったが,生理的条件ではプロトン濃度が低いため,ナトリウムポンプとして働くことになる29).レチナールシッフ塩基部位は光によるスイッチであり,第一ゲートとして働く.この場合,プロトンが取り込まれるとシッフ塩基に結合して図4のような反応サイクルを示す一方,ナトリウムイオンが取り込まれると発色団より細胞外側寄りの内部に結合する.このとき,もう一つのゲート(第二ゲート)が閉じていることが必須であり,このため海水中のナトリウムイオンが結合することはない.このように,通常のパナマ運河モデルでは説明できない光駆動ナトリウムポンプであるが,パナマ運河と同様,二つのゲートを巧みに制御することで能動輸送を実現する29).能動輸送に新しい概念をもたらした光駆動ナトリウムポンプであるが,オプトジェネティクスのツールとしても期待されている.なお,光駆動ナトリウムポンプや後述する新たなロドプシンの発見と解析は,助教・准教授としてラボに在籍した井上圭一博士(現東大物性研准教授)を中心に行った仕事である.
プロトンの濃度勾配がATP産生に使われることは細菌からヒトまで共通であり,このため生物は呼吸鎖の膜タンパク質群に代表されるさまざまなプロトン汲出し装置を創り上げてきた.その方向はすべて外向きであり,内向きプロトンポンプはATP合成酵素と競合するため,生物にとって自殺行為である.私は外向きプロトンポンプのメカニズムを深く理解するためにも,自然界に存在しない内向きプロトンポンプを創成したいと考え,試みてきた.その根拠となったのは,BRのAsp85をThrに置換したことによる内向きプロトンポンプから内向きクロライドポンプへの機能転換の成功例である33).1995年に達成した1アミノ酸置換による機能転換はロドプシンにおける最初の機能転換であり,分野を超えて大きなインパクトを与える結果となった.
内向きプロトンポンプの創成については,BRなどの外向きプロトンポンプを鋳型にしては実現しなかったが,意外なところから突破口が開かれた.大学院生だった川鍋陽博士(現香川大講師)が藍藻の光センサーであるAnabaena sensory rhodopsin(ASR)の赤外分光解析を行う過程で,ある部位のアスパラギン酸をグルタミン酸に変異させるとプロトン化の信号が増大するのを見いだした.ASRにはイオン輸送性がないのだが,彼はこの変異体がイオンポンプとして働くのではと考え調べてみたところ内向きプロトン輸送を見いだしたのである34).これまで細胞小器官の酸性化はBRなどの外向きプロトンポンプにより実現できるが,アルカリ化を実現する光遺伝学ツールは初めてであり,実際に初期エンドソーム内のpHを制御することでAMPA受容体のエンドサイトーシスを光制御する新規ツールとしてシナプス可塑性の研究に期待されている35).
内向きプロトンポンプの創製は基礎研究においても応用研究においても重要なものであったが,それより驚きであったのは,内向きプロトンポンプ機能を持つロドプシンが自然から見つかったことである.我々はゲノム科学の進展により次々に報告されるロドプシンの配列を検討し,面白そうなタンパク質の物性を検討してきたわけであるが,最初に着目するのは第三ヘリックスのモチーフである.配列が報告されるすべての微生物ロドプシンの9割以上が海洋性細菌のプロトンポンプでDTEモチーフを持つ中,Parvularcula oceaniという海洋性細菌のロドプシン(PoXeR)がASRとよく似た配列(一致度51%)を持っており,大腸菌に発現させると,内向きプロトンポンプ機能を示したのである36).哺乳類細胞に発現させて行った電気生理学的解析でも内向きプロトンポンプ活性を確認した.
PoXeRとASRのモチーフはDTLとDTSであるが,配列上の特徴は第七ヘリックスにある.BRではD85とともにD212がプロトン化シッフ塩基の対イオンとして存在し,これはクロライドポンプやナトリウムポンプ,センサー,チャネルなどに高度に保存されている.PoXeRとASRの場合,レチナールが結合するリシンから細胞外側寄りに1ターン離れたこの残基がプロリンであることが特徴である(図5).一方,細胞質側に2ターン離れた残基がアスパラギン酸であり,赤外分光解析の結果,この残基がプロトンアクセプターとして機能することがわかった36).興味深いことに,ASRもこの部位にアスパラギン酸を持つのであるが,グルタミン酸に変異させたときに内向きプロトンポンプとなったのである34).
内向きプロトンポンプのメカニズムを分光解析した結果を外向きプロトンポンプと比較して図8に示す.初期過程はすべての微生物ロドプシンと同様,all-trans型から13-cis型への光異性化反応であるが,続いて起こるプロトン移動の方向が逆向きになる.外向きであれば細胞外側へ,内向きであれば細胞質側へとプロトン移動が起こる.方向性を決める鍵になるのがシッフ塩基の正電荷を安定化させる対イオンである.BRにおけるD85とD212のように,ほとんどのロドプシンで二つの負電荷が存在するが,内向きプロトンポンプでは配列上の特徴として一つだけである.プロトン移動が起こるには,ドナーのpKaが下がると同時にアクセプターのpKaが上がる必要がある.PoXeRのように対イオンを一つしか持たない場合,pKa上昇が起こらず,対イオンはプロトンを受容できないものと考えられる.その後,いずれの場合も反対側からプロトンを受け取ることで一方向輸送が実現するが,13-cis型からall-trans型へ熱異性化する外向きプロトンポンプと異なり,内向きプロトンポンプではC13=C14結合ではなくC15=N結合が熱異性化してプロトンを受け取り,最後にC13=C14結合とC15=N結合が同時に異性化する「自転車ペダル」的熱異性化反応により光反応が完結する36).このような複雑な異性化反応はPoXeRの光反応サイクルの遅さ(1分程度かかる)を説明するものであるが,これではポンプの効率として悪すぎるように思える.さらに詳細な解析の結果,内向きプロトンポンプは図8に示す異性化過程に加えて,13-cis型からall-trans型へと異性化する速い反応成分も含まれることがわかった37).内向きプロトンポンプの構造は外向きプロトンポンプと非常によく似ており38),類似の構造からレチナール近傍のわずかな違いが,プロトンの方向性を逆転させたポンプを生み出していることが明らかになった.
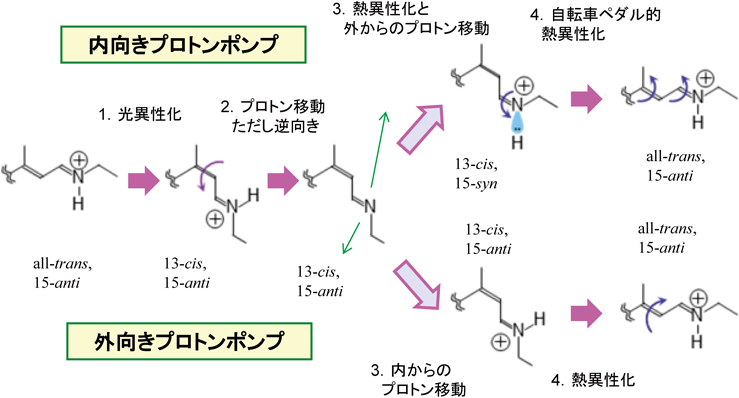
外向きポンプと内向きポンプで初期反応は共通であるが,プロトン移動の向きが逆になる.
ロドプシンはタイプ1と呼ばれる微生物ロドプシンと,タイプ2と呼ばれる動物ロドプシンに分類される.これまでに紹介したものも含め,現在,知られている機能を図9に示した.動物ロドプシンのほぼすべてがGPCRとして水溶性タンパク質(Gタンパク質)に情報を伝える光センサーであるのに対して,微生物ロドプシンの機能は実に多様である.すでに紹介したセンサリーロドプシンI,センサリーロドプシンIIが情報を伝えるのは膜タンパク質であるが,ASRは水溶性の伝達タンパク質に情報を伝える.また,陽イオンチャネルのチャネルロドプシンを変異により陰イオンチャネルに機能転換した報告の直後,2015年には天然の陰イオンチャネルが発見された39).激しい競争の中,結晶構造も決定され,構造基盤に立脚したメカニズム研究が進んでいる40, 41).一方,2014年には光でcyclic GMP(cGMP)濃度を上昇させる酵素ロドプシン(Rh-GC)が発見され42),ロドプシンの機能はついに酵素反応にまで及んだ.我々のグループは光でcAMPやcGMP濃度を低下させる酵素ロドプシン(Rh-PDE)を2017年に発見したが43),酵素ロドプシンは新しいタイプのオプトジェネティクスツールとして大きな可能性を持っている.
このようにここ数年でロドプシンの機能が驚くほどの拡がりを示す中,さらなる驚きが2018年に待っていた.上述した微生物ロドプシンは,多様な機能にもかかわらず,アミノ酸配列の相同性を持ち,系統樹を書くことができる.ところが微生物ロドプシンとも動物ロドプシンとも配列相同性を持たないロドプシン様タンパク質分子が見つかったのである.イスラエルのBéjàグループは環境中のDNA断片をもとに単離・培養が困難な微生物の遺伝子を解析するメタゲノム解析のエキスパートであり,実際にPRを発見したことで有名である18).彼らは最近,ロボットを使って環境中の遺伝子断片を個別に導入した大腸菌にレチナールを加えて色がつくかどうかでロドプシンの存在を調べる手法(機能性メタゲノム解析法)を開発し,イスラエルのガリラヤ湖に存在する遺伝子群を調べたところ,紫色を示す遺伝子産物を見いだした.ところが,このタンパク質のアミノ酸配列は微生物型とも動物型ともまったく異なっていたのである.
Béjà教授から物性解析に関する共同研究の依頼があったとき,私は絶対に何かの間違いと思っていたのであるが,配列相同性を持たないにもかかわらず,調べれば調べるほど微生物ロドプシンとよく似ていた.このタンパク質はall-trans型レチナールを持ち,光を吸収すると13-cis型に異性化した後,プロトン移動が起こって大きな構造変化が起こる光反応サイクルを示したのであるが44),それは図4に示す微生物ロドプシンの光反応性そのものである.また配列をよくみると,第七ヘリックスにリシン,第三ヘリックスにグルタミン酸があり,それぞれレチナール結合と対イオンとして働くことがわかった.我々はタイプ3のロドプシンともいえるこのタンパク質に太陽を意味するギリシャ語からヘリオロドプシンと名づけたのであるが(図10),いったんロドプシンであることが判明すると真正細菌,古細菌,ウイルス,藻類・菌類などの真核生物から500種類以上のヘリオロドプシンが見つかった.ヘリオロドプシンにイオン輸送能はなく,光反応が遅いことからセンサーであると考えているが,機能は未解明である.興味深いことに,ロドプシンはすべてN末端側が細胞外に向くが,ヘリオロドプシンは膜に対するトポロジーが逆転してN末端を細胞質に面しており(図10)44),その機能との相関が予想される.配列検索からは到達するのが不可能であったヘリオロドプシンの発見は,さらなるロドプシンワールドの可能性,発展性を示唆するものかもしれない.
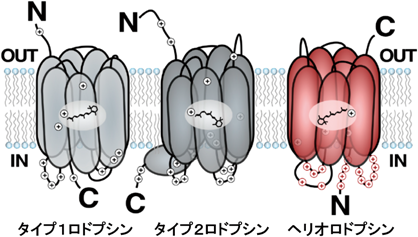
タイプ1と呼ばれる微生物ロドプシン,タイプ2と呼ばれる動物ロドプシンと異なり,ヘリオロドプシンは膜に対する配向が逆転している.ロドプシン分野の慣習と異なり,この図は細胞質側を下に書いていることに注意.文献44の図を改変.
私はロドプシン研究により学位を取得した直後のポスドク時代(1990年代初頭)や名工大に移ったころ(2000年代初頭),「まだロドプシンを研究しているのですか?」といった質問を受けることがあった.興味深いことに,最近ではそのような問いはまったく受けず,ロドプシン研究の面白みに関する感想をいただくことの方が多い.次々に登場する新しいロドプシンのメカニズムという基礎研究とオプトジェネティクスという応用研究の相乗効果により,ロドプシン研究は大きな注目を集めているといえるだろう.
本稿では我々の研究を中心に微生物ロドプシンの最新の知見について紹介した.我々は片山耕大博士(現名工大助教)を中心に動物ロドプシンである霊長類の色覚視物質の赤外分光研究45, 46)も行っており,これは現在も色覚視物質に関する世界で唯一の構造研究である.光を生物機能につなげるロドプシンを理解するには,量子力学が支配するフェムト(10−15)秒から進化をもたらすペタ(1015)秒まで,時間軸で30桁に及ぶ事象が対象となり,そのためにはさまざまな実験的手法,計算科学的手法が期待される.実際に我々が最近行った波長制御に関する研究では,構造に基づいた量子化学計算47)という「正統的な」計算科学的解析に加えて,機械学習を用いた手法48)も試みている.ロドプシンというこの魅力的な分子をより深く理解するため,オリジナルな方法論を持つ研究者のさらなる参入が期待される.
1) 神取秀樹(2015)光受容タンパク質によるエネルギー変換機構.生物物理,55, 291–298.
2) 秋田総理,菅 倫寛,沈 建仁(2017)光合成水分解・酸素発生反応の構造基盤.生化学,89, 699–709.
3) Ernst, O.P., Lodowski, D.T., Elstner, M., Hegemann, P., Brown, L.S., & Kandori, H. (2014) Microbial and animal rhodopsins: structures, functions, and molecular mechanisms. Chem. Rev., 114, 126–163.
4) 寺北明久,七田芳則(2005)ロドプシンの分子設計とその多様性,生物物理,45, 302–307.
5) Kandori, H., Matuoka, S., Shichida, Y., Yoshizawa, T., Ito, M., Tsukida, K., Balogh-Nair, V., & Nakanishi, K. (1989) Mechanism of isomerization of rhodopsin studied by use of 11-cis-locked rhodopsin analogues excited with a picosecond laser pulse. Biochemistry, 28, 6460–6467.
6) Kandori, H., Mizukami, T., Okada, T., Imamoto, Y., Fukada, Y., Shichida, Y., & Yoshizawa, T. (1990) Bathoiodopsin, a primary intermediate of iodopsin at physiological temperature. Proc. Natl. Acad. Sci. USA, 87, 8908–8912.
7) Oesterhelt, D. & Stoeckenius, W. (1971) Rhodopsin-like protein from the purple membrane of Halobacterium halobium. Nat. New Biol., 233, 149–152.
8) Henderson, R. & Unwin, N. (1975) Three-dimensional model of purple membrane obtained by electron microscopy. Nature, 257, 28–32.
9) 神取秀樹(2018)なぜロドプシンは細胞質側が上なのか?Richard Henderson氏のノーベル賞に寄せて.生物物理,58, 103–105.
10) 神山 勉(2010)光駆動イオンポンプの作動原理.生物物理,50, 222–227.
11) Nango, E., Royant, A., Kubo, M., Nakane, T., Wickstrand, C., Kimura, T., Tanaka, T., Tono, K., Song, C., Tanaka, R., et al. (2016) A three-dimensional movie of structural changes in bacteriorhodopsin. Science, 354, 1552–1557.
12) Kandori, H. (2000) Role of internal water molecules in bacteriorhodopsin. Biochim. Biophys. Acta, 1460, 177–191.
13) 神取秀樹(2011)ロドプシンの分子科学.Mol. Sci., 5, A0043.
14) Matsuno-Yagi, A. & Mukohata, Y. (1977) Two possible roles of bacteriorhodopsin; A comparative study of strains of Halobacterium halobium differing in pigmentation. Biochem. Biophys. Res. Commun., 78, 237–243.
15) Bogomolni, R.A. & Spudich, J.L. (1982) Identification of a third rhodopsin-like pigment in phototactic Halobacterium halobium. Proc. Natl. Acad. Sci. USA, 79, 6250–6254.
16) Takahashi, T., Tomioka, H., Kamo, N., & Kobatake, Y. (1985) A photosystem other than PS370 also mediates the negative phototaxis of Halobacterium halobium. FEMS Microbiol. Lett., 28, 161–164.
17) Sudo, Y., Iwamoto, M., Shimono, K., Sumi, M., & Kamo, N. (2001) Photo-induced proton transport of pharaonis phoborhodopsin (sensory rhodopsin II) is ceased by association with the transducer. Biophys. J., 80, 916–922.
18) Béjà, O., Aravind, L., Koonin, E.V., Suzuki, M.T., Hadd, A., Nguyen, L.P., Jovanovich, S.B., Gates, C.M., Feldman, R.A., Spudich, J.L., et al. (2000) Bacterial rhodopsin: Evidence for a new type of phototrophy in the sea. Science, 289, 1902–1906.
19) Nagel, G., Ollig, D., Fuhrmann, M., Kateriya, S., Musti, A.M., Bamberg, E., & Hegemann, P. (2002) Channelrhodopsin-1: A light-gated proton channel in green algae. Science, 296, 2395–2398.
20) Boyden, E.S., Zhang, F., Bamberg, E., Nagel, G., & Deisseroth, K. (2005) Millisecond-timescale, genetically targeted optical control of neural activity. Nat. Neurosci., 8, 1263–1268.
21) Ishizuka, T., Kakuda, M., Araki, R., & Yawo, H. (2006) Kinetic evaluation of photosensitivity in genetically engineered neurons expressing green algae light-gated channels. Neurosci. Res., 54, 85–94.
22) Deisseroth, K. (2011) Optogenetics. Nat. Methods, 8, 26–29.
23) Zhang, F., Wang, L.P., Brauner, M., Liewald, J.F., Kay, K., Watzke, N., Wood, P.G., Bamberg, E., Nagel, G., Gottschalk, A., et al. (2007) Multimodal fast optical interrogation of neural circuitry. Nature, 446, 633–639.
24) Chow, B.Y., Han, X., Dobry, A.S., Qian, X., Chuong, A.S., Li, M., Henninger, M.A., Belfort, G.M., Lin, Y., Monahan, P.E., et al. (2010) High-performance genetically targetable optical neural silencing by light-driven proton pumps. Nature, 463, 98–102.
25) 神取秀樹他(2013)オプトジェネティクス—光工学と遺伝学による行動抑制技術の最前線—.NTS.
26) Yawo, H., Kandori, H., & Koizumi, A. eds. (2015) Optogenetics, Light-Sensing Proteins and Their Applications. Springer Japan.
27) Inoue, K., Ono, H., Abe-Yoshizumi, R., Yoshizawa, S., Ito, H., Kogure, K., & Kandori, H. (2013) A light-driven sodium ion pump in marine bacteria. Nat. Commun., 4, 1678.
28) Kandori, H. (2015) Ion-pumping microbial rhodopsins. Front. Mol. Biosci., 2, 52.
29) Kandori, H., Inoue, K., & Tsunoda, S. (2018) Light-driven sodium-pumping rhodopsin: A new concept of active transport. Chem. Rev., 118, 10646–10658.
30) Kato, Y., Inoue, K., & Kandori, H. (2015) Kinetic analysis of H(+)-Na(+) selectivity in a light-driven Na(+)-pumping rhodopsin. J. Phys. Chem. Lett., 6, 5111–5115.
31) Kato, H.E., Inoue, K., Abe-Yoshizumi, R., Kato, Y., Ono, H., Konno, M., Hososhima, S., Ishizuka, T., Hoque, M.R., Kunitomo, H., et al. (2015) Structural basis for Na(+) transport mechanism by a light-driven Na(+) pump. Nature, 521, 48–53.
32) Konno, M., Kato, Y., Kato, H.E., Inoue, K., Nureki, O., & Kandori, H. (2016) Mutant of a light-driven sodium ion pump can transport cesium ions. J. Phys. Chem. Lett., 7, 51–55.
33) Sasaki, J., Brown, L.S., Chon, Y.-S., Kandori, H., Maeda, A., Needleman, R., & Lanyi, J.K. (1995) Conversion of bacteriorhodopsin into a chloride ion pump. Science, 269, 73–75.
34) Kawanabe, A., Furutani, Y., Jung, K.H., & Kandori, H. (2009) Engineering an inward proton transport from a bacterial sensor rhodopsin. J. Am. Chem. Soc., 131, 16439–16444.
35) Kakegawa, W., Katoh, A., Narumi, S., Miura, E., Motohashi, J., Takahashi, A., Kohda, K., Fukazawa, Y., Yuzaki, M., & Matsuda, S. (2018) Optogenetic control of synaptic AMPA receptor endocytosis reveals roles of LTD in motor learning. Neuron, 99, 985–998.
36) Inoue, K., Ito, S., Kato, Y., Nomura, Y., Shibata, M., Uchihashi, T., Tsunoda, S.P., & Kandori, H. (2016) A natural light-driven inward proton pump. Nat. Commun., 7, 13415.
37) Inoue, K., Tahara, S., Kato, Y., Takeuchi, S., Tahara, T., & Kandori, H. (2018) Spectroscopic study of proton-transfer mechanism of inward proton-pump rhodopsin, parvularcula oceani xenorhodopsin. J. Phys. Chem. B, 122, 6453–6461.
38) Shevchenko, V., Mager, T., Kovalev, K., Polovinkin, V., Alekseev, A., Juettner, J., Chizhov, I., Bamann, C., Vavourakis, C., Ghai, R., et al. (2017) Inward H+ pump xenorhodopsin: Mechanism and alternative optogenetic approach. Sci. Adv., 3, e1603187.
39) Govorunova, E.G., Sineshchekov, O.A., Janz, R., Liu, X., & Spudich, J.L. (2015) NEUROSCIENCE. Natural light-gated anion channels: A family of microbial rhodopsins for advanced optogenetics. Science, 349, 647–650.
40) Kim, Y.S., Kato, H.E., Yamashita, K., Ito, S., Inoue, K., Ramakrishnan, C., Fenno, L.E., Evans, K.E., Paggi, J.M., Dror, R.O., et al. (2018) Crystal structure of the natural anion-conducting channelrhodopsin GtACR1. Nature, 561, 343–348.
41) Kato, H.E., Kim, Y.S., Paggi, J.M., Evans, K.E., Allen, W.E., Richardson, C., Inoue, K., Ito, S., Ramakrishnan, C., Fenno, L.E., et al. (2018) Structural mechanisms of selectivity and gating in anion channelrhodopsins. Nature, 561, 349–354.
42) Avelar, G.M., Schumacher, R.I., Zaini, P.A., Leonard, G., Richards, T.A., & Gomes, S.L. (2014) A rhodopsin-guanylyl cyclase gene fusion functions in visual perception in a fungus. Curr. Biol., 24, 1234–1240.
43) Yoshida, K., Tsunoda, S.P., Brown, L.S., & Kandori, H. (2017) A unique choanoflagellate enzyme rhodopsin exhibits light-dependent cyclic nucleotide phosphodiesterase activity. J. Biol. Chem., 292, 7531–7541.
44) Pushkarev, A., Inoue, K., Larom, S., Flores-Uribe, J., Singh, M., Konno, M., Tomida, S., Ito, S., Nakamura, R., Tsunoda, S.P., et al. (2018) A distinct abundant group of microbial rhodopsins discovered using functional metagenomics. Nature, 558, 595–599.
45) Katayama, K., Furutani, Y., Imai, H., & Kandori, H. (2010) An FTIR study of monkey green- and red-sensitive visual pigments. Angew. Chem. Int. Ed., 49, 891–894.
46) Katayama, K., Nonaka, Y., Tsutsui, K., Imai, H., & Kandori, H. (2017) Spectral tuning mechanism of primate blue-sensitive visual pigment elucidated by FTIR spectroscopy. Sci. Rep., 7, 4904.
47) Inoue, K., Marín, M.D.C., Tomida, S., Nakamura, R., Nakajima, Y., Olivucci, M., & Kandori, H. (2019) Red-shifting mutation of light-driven sodium-pump rhodopsin. Nat. Commun., 10, 1993.
48) Karasuyama, M., Inoue, K., Nakamura, R., Kandori, H., & Takeuchi, I. (2018) Understanding colour tuning rules and predicting absorption wavelengths of microbial rhodopsins by data-driven machine-learning approach. Sci. Rep., 8, 15580.

名古屋工業大学大学院工学研究科教授.同オプトバイオテクノロジー研究センターセンター長.理学博士.
1960年愛知県生.84年京都大学理学部卒業(物理学教室),89年同大学院理学研究科博士課程修了(生物物理学教室).分子研ポスドク,理研ポスドク,京大院理助手,講師を経て,2001年名古屋工業大学助教授,03年より教授.
研究テーマと抱負光応答性タンパク質のメカニズム研究.特に,赤外分光法を用いたロドプシンの光情報変換・エネルギー変換機構の解明と機能開拓.
ウェブサイトhttp://www.ach.nitech.ac.jp/~physchem/kandori/index_j.html
趣味サッカー(1986年にグアダラハラでブラジルvsフランス戦を観戦したのが自慢).

This page was created on 2019-07-11T10:39:53.162+09:00
This page was last modified on 2019-08-08T13:29:03.000+09:00
このサイトは(株)国際文献社によって運用されています。