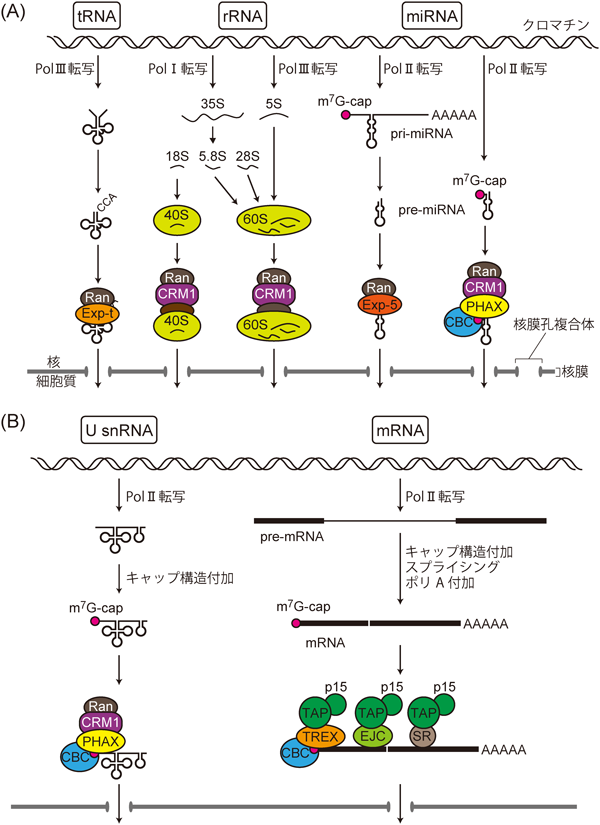
DNAの塩基配列にコードされている遺伝情報は,転写によってRNAの塩基配列へと変換される.その後RNAはプロセシングされ,活性を持つRNA分子となったりタンパク質へ翻訳されたりする.真核細胞は転写と翻訳の場が核膜によって隔てられており,多くのRNAが核膜に穿たれた核膜孔を通って細胞質に現れる.RNAの細胞質への出現はRNA単独で起こるのではなく,輸送因子群によって行われる.その際messenger RNA(mRNA),transfer RNA(tRNA),ribosomal RNA(rRNA),Uridine-rich small nuclear RNA(U snRNA),microRNA前駆体(pre-miRNA)などのRNAは,その種類に対応した特異的な輸送因子群と複合体を形成して核外へ輸送される1–4)(図1).このことは,「異なる種類のRNAは異なる経路によって核外へ輸送される」と表現される.この一般的な規則は以下の先見的な研究から明らかになった.
JarmolowskiらはRNAの核外輸送を解析する手法として,アフリカツメガエルの卵母細胞へのRNA顕微注入法を用いた.これは,試験管内転写反応によって放射能標識したRNAを卵母細胞の核内へ顕微注入し,経時的に核と細胞質に存在するRNAの分配を調べる手法である.さまざまな種類のRNAがどのような機構で核から細胞質へ輸送されるのかを調べる目的で,標識RNAと一緒に非標識のRNAを大量に注入するという実験が行われた.その結果,非標識tRNAを注入すると標識tRNAの核外輸送が遅延することが明らかになった.この結果は,大量のtRNAによってtRNA輸送因子群が飽和してしまい,tRNA自身の輸送が競合的に阻害されたと解釈することができる.同様の現象は,mRNA,rRNA,およびU snRNAの輸送の場合にも観察された.一方,たとえば標識tRNAの核外輸送の場合に,非標識のmRNA,rRNA,U snRNAをそれぞれ大量に注入しても競合阻害はみられなかった.このことから,tRNAの核外輸送はそれ以外の種類のRNAとは異なる輸送因子群を利用していることが明らかになった(実際には部分的に共通の因子を利用している場合がある).tRNA以外のRNAについても同様の検証がなされ,「異なる種類のRNAは異なる経路によって輸送される」ことが一般化された4).この実験を嚆矢として,卵母細胞への顕微注入法や生化学的解析,酵母を用いた遺伝学的手法などにより多様な研究が活発に行われ,各種RNAの輸送因子群の同定とRNA輸送複合体形成機構の理解が急速に進むこととなった.
次節ではまず,一般的なmRNAの核外輸送複合体形成について概説する.同時に,U snRNAの核外輸送についてもふれる.mRNAはU snRNAと同じくRNAポリメラーゼⅡ型(RNA polymeraseII: PolII)によって転写されるだけでなく,U snRNAと類似した特徴を備えている.にも関わらず,一般的なmRNAはU snRNAとは異なる経路で核外輸送される.mRNAの核外輸送において,どのような機構によってU snRNA型の輸送経路の利用が抑制されているのだろうか.もし,その抑制機構が破綻してmRNAにU snRNA型の輸送複合体が形成された場合,mRNAにどのような問題が生じるのだろうか.
mRNA核外輸送複合体の形成機構の詳細については片平氏の項を参照されたい.
1)U snRNAの核外輸送複合体
U snRNAはPolIIによって転写されるので,新生RNA鎖の5′末端に7-メチルグアノシン型(m7G)のキャップ構造が転写開始直後に付加される.このキャップ構造がU snRNAの核外輸送に重要なシグナルとなる.これにキャップ結合タンパク質複合体(cap binding complex: CBC)が結合する.そしてCBCとRNAの両方に結合するPHAXがU snRNAのキャップ構造近傍に結合する.次いでPHAXの核外輸送シグナル(nuclear export signal: NES)を介して,核外輸送因子のCRM1がRanGTP依存的にU snRNA上にリクルートされる.こうした仕組みにより,U snRNAはCRM1に依存した機構(CRM1経路)によって核外輸送される5)(図1B).核外へ輸送されたU snRNAは細胞質で一連のプロセシングを経て,核内に輸送される.そして再輸送されたU snRNAはmRNA前駆体のスプライシング反応に関わる.
2)mRNAの核外輸送複合体形成
一般的なmRNAの核外輸送にもキャップ構造は重要な役割を果たす.mRNAもPolIIによって転写されるのでキャップ構造が付加され,これにCBCが結合する.ここまではU snRNAの場合と同じである.しかし,これ以降の輸送因子群はまったく異なっている.mRNAのCBCにはPHAXは結合せず,一般的なmRNAの核外輸送はCRM1経路ではない.その替わりにRNA結合タンパク質のALY/REFが,CBCとRNAに結合する6,7).この際ALY/REFはtranscription/export(TREX)複合体と呼ばれるタンパク質複合体の構成因子としてmRNA上にリクルートされる.このTREX複合体が,核外輸送因子のTAP/NXF1-p15ヘテロ二量体(以下,TAP-p15)をRNA上にリクルートすることにより,mRNA輸送複合体がRNAのキャップ構造近傍に形成される.このような仕組みによって,一般的なmRNAはTAP-p15に依存した機構(TAP-p15経路)で核外輸送される(図1B).
以上のようにU snRNAとmRNAの核外輸送では,RNAのキャップ構造とCBCという共通の因子が関与しているにも関わらず,最終的な輸送複合体はまったく異なる.これは,U snRNAとmRNAの何らかの相違点が識別されていることを示している.それでは,両RNAにはどのような相違点があるのだろうか.
3)イントロン配列によるmRNAとU snRNAの識別
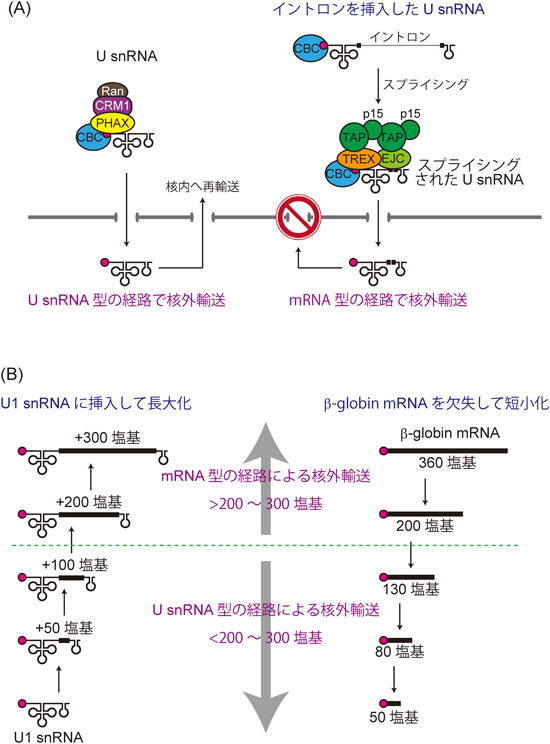
mRNAとU snRNAの相違点の一つとして,イントロン配列の有無があげられる.多くのmRNAは,イントロン配列を含んだRNA(mRNA前駆体)として転写され,mRNA前駆体は核内でスプライシングされる.一方,高等真核細胞のU snRNAにはイントロン配列がない.このことからイントロン配列の有無が,mRNAとU snRNAとを識別する特徴である可能性が考えられた.このことを検証するため,筆者らはU snRNAに人為的にイントロン配列を挿入したRNAを作製し,このRNAの卵母細胞への顕微注入実験を行った8).このRNAは核内でスプライシングされ核外へ輸送されたが,興味深いことに,スプライシングを経たU snRNAにはPHAXは結合せず,替わりにALY/REFが結合していることがわかった.このRNAの核外輸送はU snRNAとしてではなく,mRNA型のTAP-p15経路によって行われていた(図2A).以上の結果は,イントロン配列を持つということが,RNAがmRNAとして認識されるための一つの目印であることを示している8).
特筆すべきことに,PHAXはスプライシングされたU snRNAに結合していないばかりか,スプライシングを受ける前のイントロン配列を保持したU snRNAにも結合していなかった.この結果は,スプライシング反応によってRNA上のPHAXが除かれたのではなく,スプライシング反応前にすでにPHAXのRNA上へのリクルートが阻害されていたことを意味する.イントロン配列を持つこと自体に,RNAをU snRNAではないものとして認識させる効果があるようだ8).イントロン配列を持つRNA上へのPHAXリクルートの阻害の仕組みについては後に考察する.
もう一つここで言及しておきたいことは,スプライシングされて輸送されたU snRNAの運命である.上述したが,通常U snRNAは核外へ輸送後,核内へ再輸送される.しかし,スプライシングされ細胞質へ輸送されたU snRNAは,U snRNAと同じ塩基配列を持つにも関わらず,核内へ再輸送されなくなった(図2A).この結果は,核外輸送因子群は輸送を行うだけではなく,輸送後のRNAの局在にも影響を与えることを示している8).これは,正常な遺伝子発現にはRNAが適切な輸送経路を選択することが重要であることを如実に示す例である.
4)RNAの長さに応じたmRNAとU snRNAの識別
mRNAの中には,もともとイントロン配列を持たないmRNAも多く存在しており,そのようなRNAもTAP-p15経路によって核外へ輸送される.したがって,イントロン配列以外にもU snRNAとmRNAとを識別するための相違点があるはずである.その一つとして,次に筆者らはRNAの長さに注目した.U snRNAの長さは200塩基以下である一方,mRNAの平均長は約2000塩基であり,両者にはRNAの長さに大きな開きがあるからである.U snRNAとmRNAの識別にRNAの長さが果たす役割を検証するため,以下の顕微注入実験を行った.
イントロン配列を持たないmRNAを人為的に欠失して,段階的に短くしたさまざまな長さのmRNAを作製した.これらのRNAを核内に顕微注入すると,ある程度の長さを持つRNAにはALY/REFが結合してmRNA型のTAP-p15経路で輸送された.しかし短くなったmRNAにはALY/REFが結合せず,替わりにPHAXが結合してU snRNA型のCRM1経路で輸送されることがわかった.これとは対照的に,U snRNAに強い高次構造を形成しないRNA配列を挿入して長くすると,そのRNAにはPHAXが結合せず,ALY/REFが結合してTAP-p15経路で輸送された.以上の実験でCRM1経路とTAP-p15経路の切り替わりが起こるRNAの長さは,どちらの実験の場合も200から300塩基であった8,9)(図2B).以上の結果から,細胞にはRNAの長さを測って,そのRNAの核外輸送経路を仕分ける因子が存在することが想定された.
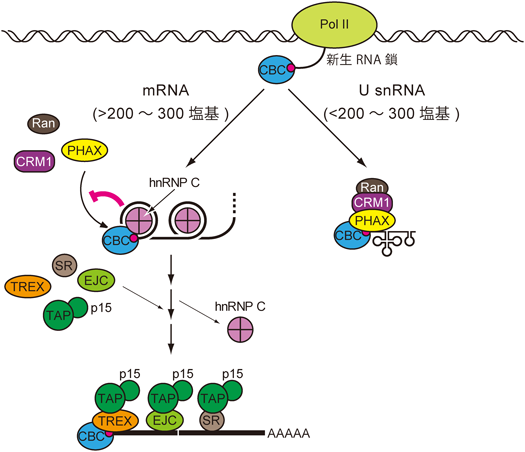
筆者らは次に,RNAの長さを測る因子を生化学的手法によりHeLa細胞の核抽出液中に探索し,核内に大量に存在するRNA結合タンパク質であるhnRNP C1/C2ヘテロ四量体であることを明らかにした10).hnRNP Cヘテロ四量体は,230から240塩基のRNAに安定に結合するという報告11)があり,この長さはちょうど上述のCRM1経路とTAP-p15経路の切り替わりが起こる境界と符合する.また,hnRNP CはCBCと相互作用することもわかった.さらに,HeLa細胞においてhnRNP CをRNA干渉法でノックダウンすると,mRNAにPHAXがリクルートされてくるようになった.この結果は,mRNA上にCRM1経路が誘導されるのを防いでいる分子がhnRNP Cであることを示唆している.また,hnRNP Cノックダウンによって,mRNAの核外輸送が遅延した(遅延を引き起こす仕組みは興味深いが現時点では明らかでない).この遅延は,hnRNP CとともにPHAXを同時にノックダウンすることによって回復した.したがってmRNAにU snRNA型の輸送複合体が形成されることは,mRNAの役割を発揮するには不適切であることを示している10).以上の結果から,hnRNP CはRNAの長さを測ることにより,mRNAとU snRNAとを仕分け,適切なmRNA輸送複合体形成を保証する役割を担っていることが明らかになった.hnRNP Cが両RNAを仕分ける仕組みとして,筆者らは以下のモデルを提唱した10).
PolIIによる転写開始直後には,新生RNA鎖の5′末端にキャップ構造が付加され,これにCBCが結合する.この段階では,新生RNA鎖がU snRNAとmRNAのどちらのRNAとして輸送されるかは未定である.RNAの長さが200塩基以下の段階で転写が終結すると,hnRNP Cの四量体は安定にはRNAに結合できない.したがってPHAXがCBCとRNAの両方に結合でき,RNAのキャップ構造近傍にU snRNA核外輸送複合体が形成され,CRM1経路で輸送される.一方,PolIIによる転写が200から300塩基以上継続して行われると,hnRNP CがRNAに安定に結合するようになる.このときhnRNP CはCBCにも結合できるので,PHAXのCBC結合は競合的に阻害される.これによりmRNAにはU snRNA型のCRM1経路は誘導されない.さらに転写反応が進行していく過程で,ALY/REFを含んだTREX複合体がhnRNP Cと置き換わる.その結果,RNAのキャップ構造近傍にmRNA核外輸送複合体が形成され,TAP-p15経路で輸送される(図3).キャップ構造近傍におけるhnRNP CからTREX複合体への置換の分子機構については明らかでないが,何らかのRNAヘリカーゼが関与しているのかもしれない.
前項で,人為的にイントロン配列を挿入することで,スプライシングされたU snRNAの輸送経路がmRNA型に切り替わった実験結果を紹介した.この切り替わりの要因として,まず第一にイントロン内のスプライシングのための塩基配列の効果が考えられる.そのような配列には,mRNAに特異的なRNA結合因子であるスプライシング因子群が集結してくるからである.ただし長さに応じたhnRNP Cによる識別を考慮すると,もう一つの可能性が考えられる.上記実験で挿入されたイントロン配列の長さは150塩基ほどであり,RNAの長さが切り替わりをある程度引き起こしているのかもしれない.スプライシングされたU snRNAの輸送経路の切り替わりが,イントロン内の配列によるものかRNAの長さによるものかを区別するためには,短いイントロン配列を用いる必要がある.しかし,RNAの長さの効果を完全に排除できるまで短くしたイントロン配列(50塩基)を挿入したU snRNAはスプライシングされなかったので,今のところ明確な答えは得られていない.
RNAの長さによってPolII転写産物が識別される機構は進化的にどれほど保存されているのだろう.予備的な結果であるが,ショウジョウバエの培養細胞でも,RNAの長さに応じてU snRNAとmRNAの輸送複合体形成が切り替わるようである.したがってショウジョウバエにも核外輸送においてRNAの長さを測る因子が働いていることが推察される.ただしhnRNP Cは後生生物に保存されているが,ショウジョウバエや線虫には明確なホモログがない.これらの生物において,どのような因子がRNA識別を行っているのか,興味深い.
5)ポリA配列によるmRNAとU snRNAの識別
一般的なmRNAのもう一つの特徴として,5′末端のキャップ構造とイントロン配列のほかに,3′末端のポリA尾部がある.ポリA尾部もU snRNAとmRNAの識別に用いられているのだろうか.筆者らはこの点についても検証した.卵母細胞への注入実験の結果,ポリA配列にはRNAをmRNA型のTAP-p15経路で輸送させる活性があることがわかった12).輸送経路の切り替わりは70塩基ほどのポリA配列でも起こり,この長さはhnRNP Cが結合するには短い.また,ポリA尾部にはポリA結合タンパク質が安定に結合する.したがって,この場合の識別にhnRNP Cが直接関与しているとは考えにくい.近年,mRNAの3′末端の切断・ポリA尾部付加反応に,TREX複合体の構成因子であるALY/REFやThoc5が関与しているという報告がある13–15).ポリA配列によるmRNA型輸送への切り替えには,TREX複合体が関与しているかもしれない.
6)CRM1経路で輸送されるmRNA
これまで説明してきたように一般的なmRNAはTAP-p15経路で輸送されるが,CRM1経路で輸送されるmRNAの知見が蓄積してきた.その代表的なものがサイクリンD1遺伝子のmRNAである16).このmRNAには核外輸送のための配列(eIF4E-sensitive element: eIF4E-SE)が存在し,この配列に翻訳開始因子のeIF4Eが結合する.eIF4Eを介してCRM1がリクルートされて,サイクリンD1 mRNAは核外輸送されることがわかった.興味深いことに,このmRNAへのTAP-p15の結合は阻害されていた16).一般的にはeIF4Eは細胞質に輸送されてきたmRNAのキャップ構造にCBCと置き換わって結合する因子である.このことを考慮すると,eIF4E-SEに結合したeIF4Eがキャップ構造にも安定に結合することで,CBC自体のキャップ結合を競合的に阻害して,ALY/REFを介したTAP-p15依存の輸送複合体形成を阻害しているのかもしれない.サイクリンD1 mRNAのような一群のmRNAの核外輸送がTAP-p15経路ではなくCRM1経路で行われる生理学的意義は不明である.
7)pre-miRNAの核外輸送との関連
mRNAと同じく,PolIIによって転写されキャップ構造やポリA尾部が付加されるRNAとして,遺伝子発現を転写後に調節するmicroRNA(miRNA)がある.miRNAはまず一次転写産物(primary miRNA: pri-miRNA)として転写される.pri-miRNAは核外輸送されず,核内でその5′側と3′側がDroshaとDGCR8から構成されるマイクロプロセッサー複合体によって切断されて,miRNA前駆体(pre-miRNA)となる.したがって,pre-miRNAはキャップ構造やポリA尾部を持たない.pre-miRNAの3′突出末端を持つ二本鎖RNAという特徴的な立体構造はエクスポーティン5(Exp-5)により認識され,RanGTP依存的にpre-miRNA輸送複合体が形成される(図1A).pre-miRNAにはキャップ構造はないので,CBCはpre-miRNAの核外輸送に直接的には関与しない.ただし,CBCはArs2と相互作用して転写中のRNAにマイクロプロセッサー複合体をリクルートすると考えられている17).マイクロプロセッサー複合体による切断の前にpri-miRNAがU snRNAやmRNAの経路で輸送されてしまわないのは,Ars2のCBC結合の効果なのかもしれない.
最近,pre-miRNAの中にExp-5依存の経路ではなく,U snRNA型のCRM1経路で核外輸送されるものが発見された18).このようなpre-miRNAはPolIIによって転写された後,マイクロプロセッサー複合体による切断を受けない.したがってキャップ構造が付加されたままであり,RNAはCBCとPHAXを介したCRM1経路によって核外輸送される(図1A).このキャップ構造を持ったpre-miRNAは200塩基以下と短いのでhnRNP Cが結合できず,核外輸送においてU snRNAとして認識されるのだろう.
3. ヒト免疫不全ウイルス1型(HIV-1)のRNAの核外輸送複合体形成の制御
一般的なmRNAにはmRNA型のTAP-p15経路だけではなく,U snRNA型のCRM1経路も誘導される可能性があるが,hnRNP CのおかげでCRM1経路の利用は抑制されることがわかった.ここでは一般的なmRNAのように,複数の核外輸送経路が誘導される可能性のあるRNAとして,HIV-1のRNAを取り上げ,このRNAの輸送経路が適切な方の一つに規定される機構について紹介する.
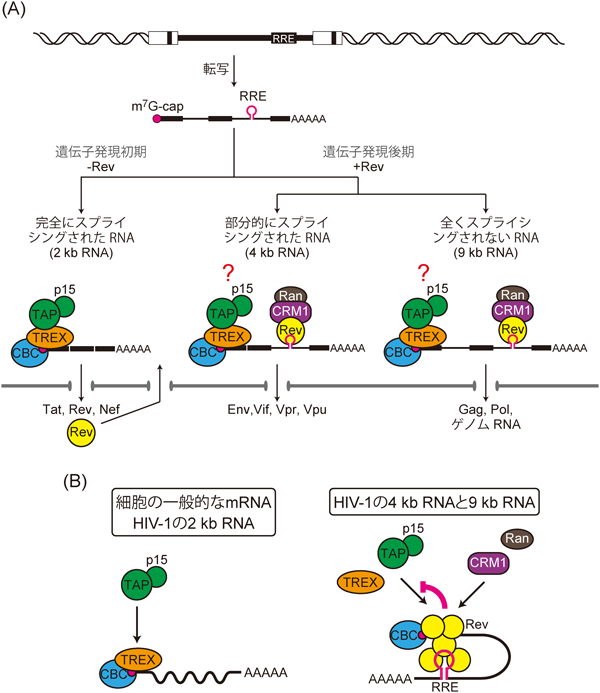
HIV-1の感染後期に発現するGag,Pol,Envなどのウイルスタンパク質は,スプライシングが不完全なウイルスRNA(長さが4000塩基と9000塩基のRNA,4 kbと9 kb)から翻訳される.そのためにはHIV-1はイントロン配列を持ったままのRNAを核外に輸送する必要がある.しかし細胞の規則としてmRNA前駆体の核外輸送は抑制されている.この細胞による監視をくぐり抜けるため,HIV-1は巧妙な戦略をとっている.それはまず,4 kb RNAと9 kb RNAのイントロン配列内に存在するRev response element(RRE)と呼ばれる配列に,ウイルスのRevタンパク質が結合する.次いでRevが,そのNESを介してCRM1をリクルートすることによって,4 kb RNAと9 kb RNAを核外輸送するというものである1)(図4A).
RevによるCRM1リクルートの活性により,4 kb RNAと9 kb RNAの核外輸送における障壁は除かれているようにみえるが,問題はそれほど単純ではない.これらのRNAの5′末端には細胞のmRNAと同じくキャップ構造が付加されているので,TREX複合体を介してTAP-p15経路も誘導される可能性がある.輸送経路が遺伝子発現に及ぼす影響を考慮すると,CRM1とTAP-p15が同一のRNAに対して同時に用いられるという状況は,HIV-1の遺伝子発現にとって問題であると考えられた(図4A).
この問題をHIV-1がどのように解決しているのかを明らかにするため,筆者らは4 kb RNAと9 kb RNAを模擬するモデルRNAを用いて解析した.その結果,RRE配列を持つRNAの核外輸送において,RevはCRM1経路を誘導する既知の活性だけではなく,ALY/REFのRNA上へのリクルートを積極的に阻害してTAP-p15経路の利用を遮断する新しい活性を持つことを見いだした.さらに,RevとCBCとの相互作用も観察され,RevはRRE配列を持つRNAのキャップ構造近傍にも結合することがわかった.以上の結果を考えると,RevはCBCに結合することで,ALY/REFを介したTAP-p15のRNA上へのリクルートを競合的に阻害しているのだろう19)(図4B).
興味深いことに,TAP-p15の過剰発現によって強制的に4 kb RNAと9 kb RNAにTAP-p15経路を誘導すると,これらのRNA量が減少し,ウイルス粒子の産生が低下した.この結果は,TAP-p15経路はウイルスの遺伝子発現に悪影響を及ぼすということ,そしてこの悪影響を回避するために,RevによるTAP-p15リクルートの阻害は重要であるということを示している19).これもまた,正常な遺伝子発現にはRNAが適切な輸送経路を選択することが重要であることを示す例である.HIV-1のRNAと上述のサイクリンD1遺伝子などの一群のmRNAはともにCRM1経路で輸送されるmRNAであるが,これらに何らかの共通の特徴があるのかもしれない.このような特徴の解明は,RNA核外輸送研究に新たな洞察を与えてくれるだろう.