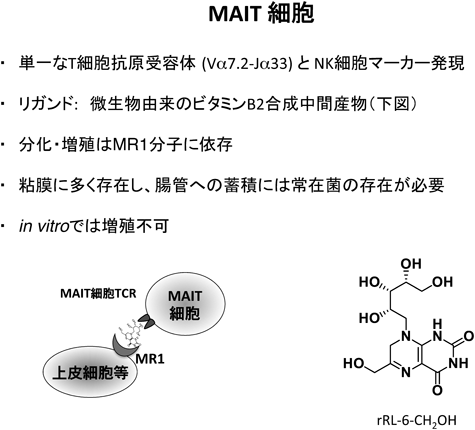
1) Salio, M., Silk, J.D., Jones, E.Y., & Cerundolo, V. (2014) Annu. Rev. Immunol., 32, 323–366.
2) Dusseaux, M., Martin, E., Serriari, N., Péguillet, I., Premel, V., Louis, D., Milder, M., Le Bourhis, L., Soudais, C., Treiner, E., & Lantz, O. (2011) Blood, 117, 1250–1259.
4) Corbett, A.J., Eckle, S.B., Birkinshaw, R.W., Liu, L., Patel, O., Mahony, J., Chen, Z., Reantragoon, R., Meehan, B., Cao, H., Williamson, N.A., Strugnell, R.A., Van Sinderen, D., Mak, J.Y., Fairlie, D.P., Kjer-Nielsen, L., Rossjohn, J., & McCluskey, J. (2014) Nature, 509, 361–365.
5) Gold, M.C., McLaren, J.E., Reistetter, J.A., Smyk-Pearson, S., Ladell, K., Swarbrick, G.M., Yu, Y.Y., Hansen, T.H., Lund, O., Nielsen, M., Gerritsen, B., Kesmir, C., Miles, J.J., Lewinsohn, D.A., Price, D.A., & Lewinsohn, D.M. (2014) J. Exp. Med., 211, 1601–1610.
6) Le Bourhis, L., Martin, E., Péguillet, I., Guihot, A., Froux, N., Coré, M., Lévy, E., Dusseaux, M., Meyssonnier, V., Premel, V., Ngo, C., Riteau, B., Duban, L., Robert, D., Huang, S., Rottman, M., Soudais, C., & Lantz, O. (2010) Nat. Immunol., 11, 701–708.
7) Georgel, P., Radosavljevic, M., Macquin, C., & Bahram, S. (2011) Mol. Immunol., 48, 769–775.
9) Miyazaki, Y., Miyake, S., Chiba, A., Lantz, O., & Yamamura, T. (2011) Int. Immunol., 23, 529–535.
10) Serriari, N.E., Eoche, M., Lamotte, L., Lion, J., Fumery, M., Marcelo, P., Chatelain, D., Barre, A., Nguyen-Khac, E., Lantz, O., Dupas, J.L., & Treiner, E. (2014) Clin. Exp. Immunol., 176, 266–274.
11) Chiba, A., Tajima, R., Tomi, C., Miyazaki, Y., Yamamura, T., & Miyake, S. (2012) Arthritis Rheum., 64, 153–161.
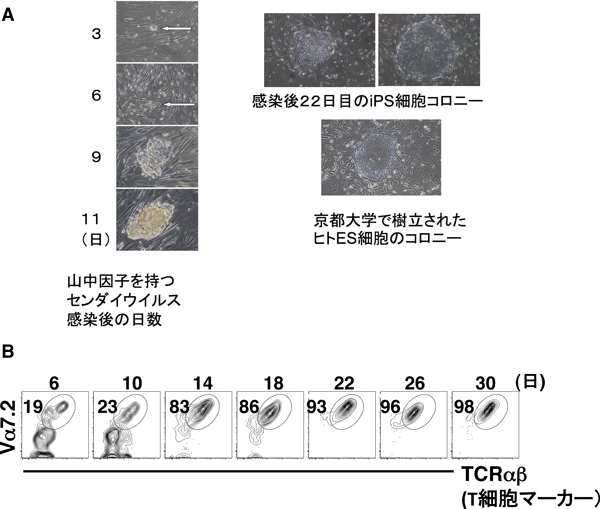
12) Wakao, H., Yoshikiyo, K., Koshimizu, U., Furukawa, T., Enomoto, K., Matsunaga, T., Tanaka, T., Yasutomi, Y., Yamada, T., Minakami, H., Tanaka, J., Oda, A., Sasaki, T., Wakao, R., Lantz, O., Udagawa, T., Sekiya, Y., Higuchi, K., Harada, N., Nishimura, K., Ohtaka, M., Nakanishi, M., & Fujita, H. (2013) Cell Stem Cell, 12, 546–558.
13) Wakao, H., Wakao, R., Sakata, S., Iwabuchi, K., Oda, A., & Fujita, H. (2008) FASEB J., 22, 2223–2231.
14) Walch, M., Dotiwala, F., Mulik, S., Thiery, J., Kirchhausen, T., Clayberger, C., Krensky, A.M., Martinvalet, D., & Lieberman, J. (2014) Cell, 157, 1309–1323.
15) Le Bourhis, L., Dusseaux, M., Bohineust, A., Bessoles, S., Martin, E., Premel, V., Coré, M., Sleurs, D., Serriari, N.E., Treiner, E., Hivroz, C., Sansonetti, P., Gougeon, M.L., Soudais, C., & Lantz, O. (2013) PLoS Pathog., 9, e1003681.