長鎖非コードRNA(long noncoding RNA: lncRNA)は,200ヌクレオチド以上の長さであり,キャップ構造やポリ(A)鎖など,mRNAと同様の構造を持つncRNAである.超高解像度タイリングアレイ解析や次世代RNAシーケンスなどの大規模トランスクリプトーム解析の結果,lncRNAはヒトにおいて1万個以上存在することが明らかとなっている.ほとんどのlncRNAは機能未知であるが,近年いくつかのlncRNAが,分化,がん化,アポトーシスや細胞老化といった細胞運命決定において,重要な役割を担っていることがわかってきた1,2).これらlncRNAの作用機構は実に多種多様である.これまでの研究により,lncRNAが,さまざまなタンパク質との相互作用を介して,転写制御,翻訳制御,核内構造体形成やタンパク質輸送などに関与することがわかってきた.
これまでに筆者らも,ANRIL(antisense non-coding RNA in the INK4 locus)がポリコーム群(Polycomb group: PcG)と結合し,INK4遺伝子座上へのPcG結合を促進することによって,CDK(cyclin-dependent kinase)インヒビターp15およびp16の転写を抑制すること,さらにANRILが細胞老化制御に関与することを報告した3).また,Changらのグループとの共同研究の結果,CDKインヒビターp21遺伝子座上流に存在するPANDA(p21 associated ncRNA DNA damage activated)が,転写因子NF-YA(nuclear transcription factor YA)と結合し,そのDNA結合を阻害することにより,アポトーシス誘導遺伝子群の転写抑制を介して,アポトーシスを抑制することが明らかとなった4).本稿では,これらANRILとPANDAによる細胞増殖およびアポトーシス制御機構について,筆者らの研究成果を中心に,最新情報をあわせて概説する.
2. INK4遺伝子座に存在するlncRNA,ANRIL
1)INK4遺伝子座
INK4遺伝子座は,ヒト染色体9p21領域に存在する.この領域は,非常にユニークな構造を持つ.およそ40 kb内に,CDKインヒビターであるp15およびp16遺伝子が存在する.さらに,p16と同じエキソンを共有するが,異なるリーディングフレームを持つARF(p53安定化因子)も存在する.p15およびp16タンパク質は,CDK4/6の活性阻害を介してがん抑制遺伝子産物RBを正に制御する.ARFタンパク質は,ユビキチンリガーゼMDM2の活性阻害を介してがん抑制遺伝子産物p53を正に制御する.したがって,INK4遺伝子座は,がん抑制において重要な領域であると考えられている.実際,多くのヒトのがん組織において,この領域の欠損,点変異および転写抑制が報告されている5,6).
2)ANRILの構造
ANRILは,フランスの家族性メラノーマ–神経系腫瘍において欠損している領域内に存在する遺伝子として,EST(expressed sequence tag)データベースを用いた解析により同定された7).ANRILは,INK4遺伝子座内のp15およびARF遺伝子間から,これらとは逆向きに転写される.ANRILのゲノムDNAは約120 kbの長さで,19個のエキソンからなる.19番目のエキソンにポリアデニル化部位を持ち,転写後ポリ(A)付加およびスプライシングを受け,約3.8 kbの長さになる.これまでに,少なくとも13個以上のANRILアイソフォームの存在が確認されている8).興味深いことに,ANRILアイソフォームの一部は,スプライシング後,環状構造をとることがわかっているが,その機能は不明である9).
3)ANRILの発現制御
SatoやWanらの解析により,ANRILは,転写因子E2F1によって転写活性化されることが報告された10,11).ANRILのプロモーター領域には,E2F結合部位がある.クロマチン免疫沈降(chromatin immunoprecipitation: ChIP)やルシフェラーゼアッセイにより,E2F1がANRILプロモーターに直接結合し,そのプロモーターを活性化することが示された.その上流シグナルとして,DNA損傷応答経路を介していることがわかっている.DNA損傷により活性化するATM(ataxia-telangiectasia mutated)依存的に,E2F1はANRILプロモーターに結合し,転写を活性化することが示された11).
筆者らは,活性型H-Ras変異体の過剰発現により,ANRILの発現量が顕著に減少することを示した3).ヒト胎児肺線維芽細胞において,過剰のRasシグナルが入ると,細胞は一過性に過増殖するが,やがてINK4遺伝子座にあるp15およびp16遺伝子の転写が活性化され,早期細胞老化が誘導される.この経路は,がん化シグナルに対する防御機構として働いている.ANRILは,p15およびp16の転写を抑制する機能を持つことから,RasシグナルによるANRILの発現抑制は,p15/p16経路を介したがん抑制において重要であると考えられる.しかしながら,RasシグナルによるANRIL発現抑制機構については,いまだ不明である.
4)ANRILによる細胞増殖制御機構
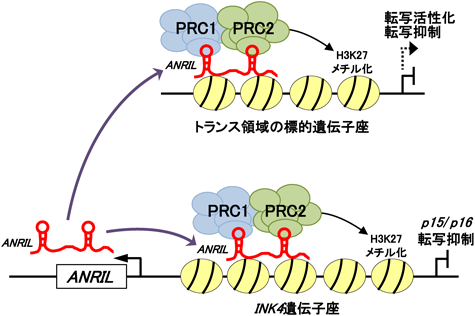
筆者らの解析の結果,ヒト胎児肺線維芽細胞WI38において,ANRILをノックダウンすると,INK4遺伝子座のp15およびp16の転写が活性化することが明らかとなった3).細胞周期のブレーキであるp15およびp16の発現量増加に伴い,ANRILをノックダウンした細胞は,増殖が顕著に抑制され,最終的に早期細胞老化が誘導された.これらの結果から,ANRILはp15およびp16遺伝子の転写を抑制し,細胞増殖,老化を抑制する機能を持つことが示唆された.ANRILによるINK4遺伝子座転写抑制機構として,筆者らやYapらは,ANRILがこの領域へのPcGリクルートメントに関与することを報告した3,12).PcGは,PRC1(Polycomb repressive complex 1)とPRC2という二つの複合体に大別される.PcGは,PRC2によるヒストンH3の27番目のリシン残基(H3K27)のメチル化と,その修飾を認識して結合するPRC1によるヒストンH2AK119のモノユビキチン化によって,標的遺伝子の転写を抑制する.以前筆者らは,PcGがヒストンH3K27のメチル化を介して,INK4遺伝子座の転写を抑制していることを報告した13).筆者らのChIPアッセイの結果,ANRILをノックダウンすると,INK4遺伝子座上のPRC2構成因子,SUZ12の結合量が減少することが示された3).さらに,SUZ12抗体を用いたRNA immunoprecipitation(RIP)アッセイの結果,ANRILがSUZ12と結合していることが示された.一方,YapらのUV cross-linking and immunoprecipitation(CLIP)アッセイの結果,クロマチン画分において,ANRILがPRC1構成因子,CBX7と結合していることが示された12).これらの結果から,ANRILは転写された後,クロマチン上にとどまり,PRC1およびPRC2と結合し,これらPcGによるINK4遺伝子座のヒストン修飾を促進させることにより,この領域の転写抑制状態を維持していることが考えられた(図1).
ANRILノックダウンによる細胞増殖の抑制は,RB経路が不活化した細胞でもみられた(未発表データ).このことは,ANRILがp15/p16以外の標的遺伝子を持つこと,つまりトランス領域の転写制御にも関与することを示唆した.最近,これを支持する研究結果が相次いで報告された.Congrainsらは,ヒトの大動脈血管平滑筋細胞において,ANRILをノックダウンすると,アテローム性動脈硬化症関連遺伝子群の発現量が変動することを示した14).興味深いことに,ANRILのエキソン1とエキソン19領域のノックダウンでは,各々変動する遺伝子群が異なっていた.ANRILには複数のアイソフォームが存在することから,各々のアイソフォームは異なる標的遺伝子あるいは機能を持つのかもしれない.一方,Holdらは,ANRILを異所的に過剰発現すると,細胞接着能,増殖能,代謝活性が亢進し,アポトーシスが抑制されることを示した8).さらにHoldらは,ANRIL過剰発現により,トランス領域(異なる染色体)の標的遺伝子座上のSUZ12およびCBX7結合量が増加することを示した.ANRIL依存的にPcGがリクルートされた標的遺伝子の転写は,抑制されるものもあれば,逆に活性化されるものもあった.ANRILは,PcGだけでなく,他の転写制御因子(この場合,転写活性化因子?)のリクルートメントにも関与しているのかもしれない.このANRILによるトランス領域の標的遺伝子発現制御には,ANRIL中にあるAlu配列が必要であった.以上のことから,ANRILはINK4遺伝子座だけでなく,トランス領域の標的遺伝子発現を制御することにより,細胞増殖,老化,アポトーシス,接着および代謝調節といった,多様な細胞機能に関与していることが考えられた(図1).今後,ANRILによるシス/トランス領域の標的遺伝子座認識機構の解明や,結合タンパク質の網羅的探索が,さらなるANRILの機能解明に必要であると考えている.
3. p21遺伝子座上流に存在するlncRNA, PANDA
1)PANDAの構造と発現
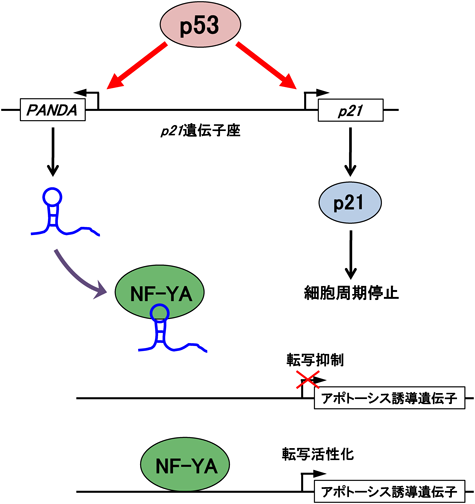
Changらと筆者らの共同研究により,細胞周期関連遺伝子群(サイクリン/CDKs/CDKインヒビター)のプロモーター領域に,216個の推定新規lncRNAが存在することが明らかとなった4).これらlncRNAの発現と,周辺の遺伝子との発現のパターンに相関性はみられなかった.この結果は,ほとんどのlncRNAがシス領域の遺伝子の転写制御に関与していないことを示唆した.これらlncRNAの中で,機能が解明されたのがPANDAである.PANDAゲノムDNAは,CDKインヒビターp21遺伝子の転写開始点の約5 kb上流に存在し,1個のエキソンからなる(図2).PANDAは,p21とは逆向きに転写され,5′末端にキャップ構造を,3′末端にポリ(A)鎖を持つ転写産物(長さ1.5 kb)となる.その発現は,DNA損傷に応答して,p53依存的に誘導された.PANDAとp21は,互いの発現には影響を及ぼさなかった.
2)PANDAによるアポトーシス制御機構
PANDAはアポトーシス制御に関与することが示された4).PANDAをノックダウンした細胞は,DNA損傷によるアポトーシスが起こりやすくなった.マイクロアレイ解析の結果,DNA損傷誘発とともにPANDAをノックダウンすると,アポトーシス誘導遺伝子群(APAF1,BIK,FAS,LRDDなど)の発現量が増加することが明らかとなった.これらの結果から,PANDAはアポトーシス誘導遺伝子群の転写を抑制することにより,アポトーシスを抑制する機能を持つことが示唆された.
p53下流のアポトーシス誘導遺伝子のプロモーターは,転写因子NF-Y複合体によって活性化される.RIPアッセイの結果,PANDAは,NF-Y複合体の構成因子であるNF-YAと結合することが示された.さらにChIPアッセイの結果,PANDAをノックダウンすると,アポトーシス誘導遺伝子群のプロモーター上のNF-YA結合量が増加することが示された.これらの結果から,DNA損傷に応答して誘導されるPANDAは,NF-YAと結合し,その標的遺伝子プロモーターへの結合を阻害することによって,アポトーシス誘導遺伝子群の転写を抑制することが考えられた(図2).
近年,lncRNAの機能解明が進むにつれ,lncRNAは多様な作用機構と細胞機能を持つことがわかってきた.本稿で注目したANRILとPANDA,両者とも転写制御に関与するが,その作用機構や細胞機能はまったく異なるものであった.この点が,lncRNA研究の興味が尽きない理由であるが,反面,一つ一つのlncRNAに対して,手探りで地道に作用機構を解明していかなければならず,解析に時間がかかる理由でもある.全機能性lncRNAの作用機構の分類や構造モチーフの同定ができる日は,はたしてやってくるのだろうか?
近年,ANRIL遺伝子座上の一塩基多型(single-nucleotide polymorphisms: SNPs)が,冠状動脈疾患,虚血性脳卒中やⅡ型糖尿病に関連することが明らかにされた.また,ANRIL遺伝子上のSNPsの一つが,ANRILの発現量減少と神経線維腫に関連することが報告された15).ANRILはがん抑制遺伝子であるp15およびp16の転写を抑制し,細胞増殖を促進する機能を持つことから,がん化に関与する可能性が考えられる.今後,ANRILやPANDAの生理機能の解明が重要な課題である,と筆者らは考えている.
謝辞Acknowledgments
本稿で紹介した研究成果は,浜松医科大学分子生物学講座北川雅敏教授,ノースカロライナ大学Yue Xiong教授,スタンフォード大学Howard Y. Chang教授をはじめ,多くの共同研究者のご指導,ご協力によるものであります.深く御礼申し上げます.
引用文献References
1) Kitagawa, M., Kitagawa, K., Kotake, Y., Niida, H., & Ohhata, T. (2013) Cell. Mol. Life Sci., 70, 4785–4794.
2) Kitagawa, M., Kotake, Y., & Ohhata, T. (2012) Curr. Drug Targets, 13, 1616–1621.
3) Kotake, Y., Nakagawa, T., Kitagawa, K., Suzuki, S., Liu, N., Kitagawa, M., & Xiong, Y. (2011) Oncogene, 30, 1956–1962.
4) Hung, T., Wang, Y., Lin, M.F., Koegel, A.K., Kotake, Y., Grant, G.D., Horlings, H.M., Shah, N., Umbricht, C., Wang, P., Kong, B., Langerod, A., Borresen-Dale, A.L., Kim, S.K., van de Vijver, M., Sukumar, S., Whitfield, M.L., Kellis, M., Xiong, Y., Wong, D.J., & Chang, H.Y. (2011) Nat. Genet., 43, 621–629.
5) Ruas, M. & Peters, G. (1998) Biochim. Biophys. Acta, 1378, F115–F177.
6) Sharpless, N.E. (2005) Mutat. Res., 576, 22–38.
7) Pasmant, E., Laurendeau, I., Heron, D., Vidaud, M., Vidaud, D., & Bieche, I. (2007) Cancer Res., 67, 3963–3969.
8) Holdt, L.M., Hoffmann, S., Sass, K., Langenberger, D., Scholz, M., Krohn, K., Finstermeier, K., Stahringer, A., Wilfert, W., Beutner, F., Gielen, S., Schuler, G., Gabel, G., Bergert, H., Bechmann, I., Stadler, P.F., Thiery, J., & Teupser, D. (2013) PLoS Genet., 9, e1003588.
9) Burd, C.E., Jeck, W.R., Liu, Y., Sanoff, H.K., Wang, Z., & Sharpless, N.E. (2010) PLoS Genet., 6, e1001233.
10) Sato, K., Nakagawa, H., Tajima, A., Yoshida, K., & Inoue, I. (2010) Oncol. Rep., 24, 701–707.
11) Wan, G., Mathur, R., Hu, X., Liu, Y., Zhang, X., Peng, G., & Lu, X. (2013) Cell. Signal., 25, 1086–1095.
12) Yap, K.L., Li, S., Munoz-Cabello, A.M., Raguz, S., Zeng, L., Mujtaba, S., Gil, J., Walsh, M.J., & Zhou, M.M. (2010) Mol. Cell, 38, 662–674.
13) Kotake, Y., Cao, R., Viatour, P., Sage, J., Zhang, Y., & Xiong, Y. (2007) Genes Dev., 21, 49–54.
14) Congrains, A., Kamide, K., Katsuya, T., Yasuda, O., Oguro, R., Yamamoto, K., Ohishi, M., & Rakugi, H. (2012) Biochem. Biophys. Res. Commun., 419, 612–616.
15) Pasmant, E., Sabbagh, A., Masliah-Planchon, J., Ortonne, N., Laurendeau, I., Melin, L., Ferkal, S., Hernandez, L., Leroy, K., Valeyrie-Allanore, L., Parfait, B., Vidaud, D., Bieche, I., Lantieri, L., Wolkenstein, P., & Vidaud, M.; NF France Network. (2011) J. Natl. Cancer Inst., 103, 1713–1722.
著者紹介Author Profile
 神武 洋二郎(こうたけ ようじろう)
神武 洋二郎(こうたけ ようじろう)近畿大学産業理工学部准教授.博士(医学).
略歴1975年福岡県に生まる.99年九州大学農学部農芸化学科卒業.2001年同大学大学院生物資源環境科学研究府修士課程修了.05年同大学大学院医学系学府博士課程修了.05年米国ノースカロライナ大学博士研究員.08年浜松医科大学助教を経て12年より現職.
研究テーマと抱負細胞老化,がん化,分化,アポトーシスなどの細胞運命決定機構の解明.特に最近では,細胞老化,がん化に関与する長鎖非コードRNAに注目し,その探索,機能解析を行っている.ヒトの疾患治療薬の分子標的やバイオマーカーとなりうる長鎖非コードRNAを見つけたい.
ウェブサイトhttp://blog.cc.fuk.kindai.ac.jp/biochem/labo/kotake/
趣味三線,アウトドア活動.
 苗村 円佳(なえむら まどか)
苗村 円佳(なえむら まどか)近畿大学大学院産業理工学研究科博士後期課程1年.
略歴1990年大阪府に生まる.13年近畿大学産業理工学部生物環境化学科卒業.15年同大学大学院産業理工学研究科修士課程修了を経て同年より現職.
研究テーマと抱負長鎖非コードRNAの機能と作用機序の解明.特に細胞周期に関与する長鎖非コードRNAに着目し,それらの機能解析を行っている.少しでも多くの長鎖非コードRNAの機能を解明したい.
趣味ドライブ,温泉巡り.
 紫 千大(むらさき ちひろ)
紫 千大(むらさき ちひろ)近畿大学大学院産業理工学研究科修士課程2年.
略歴1991年福岡県に生まる.2014年近畿大学産業理工学部生物環境化学科卒業.同年より現職.
研究テーマと抱負がん細胞に特異的に発現している長鎖非コードRNAの機能と作用機序の解明.特に細胞死に関与する長鎖非コードRNAに注目し,その作用機序と発現制御の解明を目指している.
趣味映画鑑賞,ジョギング.