生体の重要な構成成分であるタンパク質は,ゲノムDNA上に塩基配列としてコードされ,mRNAへの転写とその後の翻訳を経て生合成される.いわば生体内工場において,DNAに描かれた設計図をもとにそれを忠実に再現したタンパク質が生産されているわけである.タンパク質のアミノ酸配列はある決まった立体構造を形成するようにプログラミングされており,生産された新生ポリペプチド鎖は原則的には熱力学的な原理に基づき自発的に決められた形にフォールディングし(Anfinsenのドグマ),その機能を生体内で果たすことになる.
この工程はさまざまな方法で管理されているものの,100%の確率で間違いなく行われることは難しく,生体内では本来意図されていないタンパク質が生成することがある.その多くは生産工程の途中における修復機構や除去機構によって生体内にとどまることはないが,一部のタンパク質はその網をすり抜け生体内で存在し続ける.このようなプログラムから逸脱した結果形成されるタンパク質構造形態の一つがアミロイド線維と呼ばれるタンパク質集合体である(アミロイド線維の漢字は,医学用語として“繊維”ではなく“線維”を用いる)1).
アミロイド線維は不溶性で,生体内のさまざまな細胞組織に沈着することが報告されている.その結果起きる病気として代表的なものがアルツハイマー病,プリオン病,ハンチントン病や透析アミロイドーシスなどであり,これらは総称的に,アミロイド病やフォールディング病と呼ばれる.
アミロイド線維の立体構造は,もはや本来アミノ酸配列のプログラムが意図していたものから大きく異なった形で作動しており,その結果生体内で異常な反応を起こしていると考えられる.アミロイド線維が形成されるきっかけや進行原因を突きとめることができれば,アミロイド線維の形成を人為的に制御することができ,効果的な疾病の治療や予防への戦略考案へと結びつくだろう.
本稿では,まずアミロイドの基本的性質とともに,アミロイド線維の形成によって及ぼされると考えられる生体への不利益について述べる.次に,アミロイド異常凝集が起こる過程のうち特に初期相に注目し,これまでの報告や筆者らの結果を交え,現在考えられる分子論的な機構について考察を行いたい.アミロイド線維形成についての研究背景や今後の課題などについての概要把握の一助となれば幸いである.
2. プログラムされていない構造―アミロイド線維構造とその特徴
1)アミロイド線維の基礎的な性質
アミロイド線維研究の歴史は,19世紀半ばにVirchowらが脳から沈着物を取り出したときに遡る.当初は,沈着物がヨウ素デンプン反応を示したことから,アミロイド線維は多糖と考えられていた.その約100年後に,アミロイドの主成分がタンパク質であることが発見された.その構造に驚くほどの規則性があることに気づいたのはさらにその先で,今から約40年前のことである2).
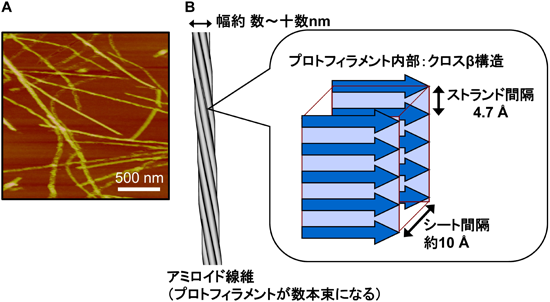
アミロイド線維を電子顕微鏡や原子間力顕微鏡で観察すると,特徴的な細長い形状が確認できる(図1A).これには枝分かれがなく,太さや長さはタンパク質の種類によって多少異なるが,通常約数~十数nmの幅と数µmの長さを持つ.またアミロイド線維にはプロトフィラメントが集合するという階層的な構造がみられ,さらにミクロレベルでは,線維軸に対してβシートが直交する“クロスβ構造”が基礎構造となっている(図1B)3).アミロイド線維構造は,特有の構造特性および機能性を持つタンパク質として存在するようにプログラムされていたポリペプチド鎖が,形成すべき立体構造を逸脱し,大々的なクロスβ構造への構造転換を経るとともに多数の分子が集まり凝集した構造と考えられる.
これまでに多種多様なタンパク質が形成する生体内凝集が確認され,解析されてきた.なかには,針状というよりはむしろ粒状の形態をした凝集体にも関わらず内部にはβシートに富んだ構造を多く含むものも確認されている.このような構造を“アミロイド様”凝集と呼んでアミロイド線維の範疇としてみなす場合も多く,実に生体内凝集に高頻度でみられる構造要素であるといえるだろう.興味深いことに,大腸菌を宿主とした発現系でしばしば観察される封入体もβシートに富んだ構造から形成されているという報告もあり4),アミロイド線維構造はポリペプチド鎖が取りうる一般的な構造として考えられている5).
2)生体内でのアミロイド構造が及ぼす影響
本来のプログラムに従って形成されたネイティブ(天然)構造である球状タンパク質では,その特有の構造や柔らかさ,ダイナミクスが酵素活性やその他の機能を発揮するのに重要な役割を果たす.しかしプログラムから逸脱したアミロイド線維では構造はもはや異なり,しかも上述のクロスβ構造が水素結合によって強固に安定化され非常に剛直な構造となっている.その結果,ネイティブ構造が有する機能が失われる(loss of function).
それだけではなく,その剛直な構造自体が細胞に対して悪影響を示すことが認められている.アミロイド線維がどのような作用を及ぼすのか,タンパク質レベルの分子論的な機構は依然十分解明されていないが,最終的にはアミロイド沈着した諸細胞が機能不全を起こすことがわかっている.具体的なメカニズムとしては,アミロイド線維が生体膜に相互作用し細胞毒性を示すことや炎症反応が進むことなどが報告されている(gain of function)6,7).アルツハイマー病の原因タンパク質であると考えられているアミロイドβタンパク質などについては,アミロイド線維が形成される以前にみられるアミロイド線維に比べ少ない会合数で凝集したオリゴマー体がさらに強い細胞毒性を有することも報告されており,2000年代に星らが10~15 nmのサイズを持つアミロスフェロイドという球状の凝集構造体がきわめて強い神経毒性を示すことを見いだし注目されている8,9).
さらに細胞毒性を考える上で非常に重要な点がアミロイド線維の自己触媒的な増殖性である.基本的な反応機構としては,アミロイド線維の末端に単量体ペプチド分子が会合することによって伸長反応が起きることが明らかとなっている.この性質が,生体内においてアミロイドの沈着が拡大していくことや,プリオン病が異常タンパク質を媒体として別の個体に感染することの分子基盤となっている可能性が考えらえる.
アミロイドが引き起こすと考えられている疾病への対策として,生成したアミロイド線維の除去とともに重要なことが,アミロイド線維の凝集を防ぐことである.筆者らはアミロイド線維が形成する初期相に焦点をあてて研究活動を行ってきた.次節ではアミロイドの基本的な形成機構について概説し,さらに最近の筆者らの研究によって得られた結果をもとに考えられる分子メカニズムを提案する.
1)フォールディングファネルの逸脱
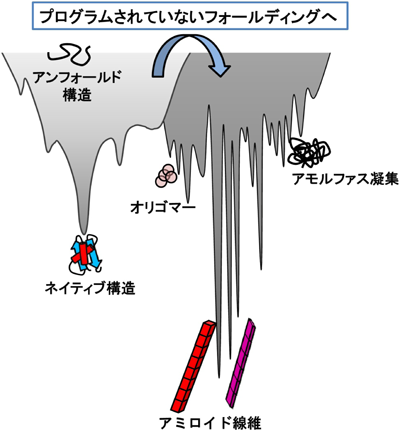
タンパク質のフォールディングを包括的に表すモデルとして,漏斗(ファネル)のような形状をしたエネルギー地形が引き合いに出されることがしばしばある.このモデルでは,縦軸でエネルギーを,横軸の広がりで構造の自由度を表す.すなわちエネルギー状態が高く自由度の高い変性構造(アンフォールド構造)はファネルの縁に存在する構造状態であり,エネルギー状態が変性状態に比べて低く,自由度も低くなったネイティブ構造はファネルの底のくぼみに位置する.しかしながら,アミロイド線維形成というタンパク質フォールディングの新たな側面を考慮しようとした場合,このモデルだけでは説明ができない.
近年,このエネルギー地形にもう一つのエネルギー地形を組み合わせることで,アミロイド線維の形成を含めた新たなモデルが提案されている(図2)10).このモデルでは,アミロイド線維形成に代表されるプログラムされていないフォールディングが起きるということは,タンパク質分子が本来のフォールディングファネルを逸脱し,通常は顕在化していない別のファネルに入り込んでしまった結果と解釈される.また,アミロイド線維では1種類のタンパク質から複数のアミロイド線維構造が形成されるという“構造多形”が確認されており,この性質が病態の複雑性を引き起こすと考えられている11,12).したがって,通常のネイティブ構造へのフォールディング地形(図2,左側薄灰着色部分のエネルギー地形)に比べ,ミスフォールディングの地形には複数のエネルギー極小状態が存在し,凹凸の多い地形であることが予想される(図2,右側濃灰着色部分のエネルギー地形).
2)アミロイド線維形成の開始を担う核形成反応
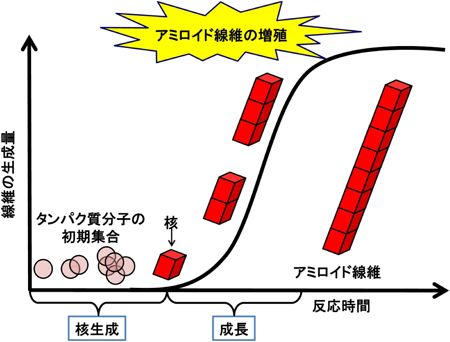
アミロイド線維の形成は,核形成とそれに続く成長の二段階で進行する.核形成相は,タンパク質分子が初期集合する過程を指しており,これによってアミロイド線維の核が形成される.その後,形成した核を鋳型として次々にペプチド鎖が凝集しアミロイド線維が増殖していくことがわかってきている.核形成は大きなエネルギー障壁に阻まれているために自発的に起こりにくく,線維形成反応全体の律速段階となっている.しかし,ひとたび核が生成すると,アミロイド線維は,自らの末端構造を鋳型として単量体が順次に結合し,成長していくことによって線維軸に沿って自己の構造を複製し,伝播させることができる.この反応様式を“核依存性伸長”という13).このように,アミロイド核の持つ構造伝播性により自己触媒的に成長・増殖し高確率で発病に至ると考えられている(図3).
3)核形成反応を加速する要因
一般に,アミロイド線維形成の初期段階には,ネイティブ構造が不安定化したり部分的な変性が起きたりすることが必要であると考えられている.さらに,タンパク質分子間の相互作用を強める因子もアミロイド線維形成反応を促進する効果があると予想される1,10,14).これらを引き起こす要因はいくつかあり,まず,前駆体タンパク質自体に施されるアミノ酸変異は,遺伝性の家族性アミロイドーシスにみられる重要かつ直接的でわかりやすい要因の一つである.たとえば,家族性アミロイドポリニューロパチーの発症メカニズムとしては,前駆体タンパク質であるトランスサイレチン中に起きている点変異が通常形成している四量体の解離を促進してしまい,その結果タンパク質の立体構造が不安定化しアミロイド線維形成反応が促進すると考えられている15).家族性アルツハイマー病の前駆体タンパク質であるアミロイドβタンパク質についても,遺伝性の強い家系から見いだされているいくつかの点変異が凝集速度を加速することが確認されている16).その他,老化に伴うアミノ酸側鎖の酸化や脱アミド化,アミノ酸のL体→D体への異性化17)といったポリペプチド鎖の化学的な修飾,金属イオンの結合,また小胞体ストレスなど生体内でのタンパク質生合成を管理する機構の不具合などが,プログラムされていないフォールディングへの引き金となっていることが報告されている14).
4)アミロイド線維を形成するポテンシャルの実体は何か?
アミロイド線維を形成するのは疾病の原因となる限られた種類のタンパク質だけなのだろうか.試験管内の実験によると,アミロイド線維を形成するポテンシャルは病気に直接関与するタンパク質だけでなくあらゆるタンパク質に本質的に備わっているようである.近年,Eisenbergらのグループは,天然由来のタンパク質のアミノ酸配列を6~8アミノ酸残基ごとに区切ることで得られたペプチド断片についてアミロイド線維形成能力を網羅的に解析し,複数の非常に安定なクロスβ構造を形成する短いアミノ酸配列領域を割り出すことに成功した18).これらの高いアミロイド線維形成能力を有するペプチドは微結晶を作ることができ,X線結晶構造解析の結果,側鎖どうしが密にパックし安定化した“steric zipper構造”を形成することが明らかとなった3).このことは,天然のタンパク質のアミノ酸配列にはアミロイド線維形成能の高いペプチド領域が含まれていることを意味する.すなわち,タンパク質のアミノ酸配列のなかにこのようなクロスβ構造を形成する能力の高い配列を持つか否か,さらにその領域がいくつ含まれるかが個々のタンパク質が潜在的に持つアミロイド性の強度を規定しているのかもしれない.
それにも関わらず正常な生体内環境ではアミロイド性が顕在化しないのは,生体内ではネイティブ構造の形成によりこれらアミロイド性の高い領域が外界から遮蔽され,分子間相互作用を獲得できないようになっているからであると考えればわかりやすい.この仮説に沿って核形成を考えると,アミロイド線維形成能の高いアミノ酸配列領域が変性反応などをきっかけとしてタンパク質分子から溶媒へ露出し,それらがさらに分子間相互作用を形成して規則的に集合する過程であると考えられる.また,先述のように,核形成相は律速段階であることが多く,アミロイド線維構造の出現ひいては疾病の発症時期を左右する重要な過程として捉えることができる.その間のタンパク質分子の集合反応がどのように進行するのかを明らかにすることは大変重要であり,タンパク質の構造形成の本質を理解することにもつながる.しかしながら現状では,アミロイド線維核構造が構築される際のタンパク質分子の初期集合過程では分光学的あるいは熱力学的なシグナルが乏しく,その実態は十分に理解されていない.
1)オリゴマー様集合体の観察例
このような現状で,核形成機構の解明のために最近特に注目が集まっているのが,オリゴマーやプロトフィブリルなど会合数がまだ比較的少ないタンパク質集合体である.これらの構造は未成熟であり,アミロイド線維にみられるような自己触媒的な増殖性もまだ発現していない.
オリゴマーの解析方法の開発は以前からさまざまなされており,特にアルツハイマー病に関与するアミロイドβタンパク質については2節で述べたように神経細胞死を引き起こす主体はオリゴマーであるという仮説が提唱されるなど,オリゴマー構造に着目した研究が進展している.これまでの注目すべきオリゴマー研究例としては,2000年代にTeplowらによって光架橋によるオリゴマー捕捉の試みが精力的に行われたことがあげられ,検出困難だったオリゴマー構造の確認および定量的解析に道筋をつけたといえる19).さらに,オリゴマー構造に特異的に結合する抗体が発見され,分布量の少ないオリゴマー体を感度よく検出することも実現した20).その他,従来のタンパク質フォールディングで用いられてきたさまざまな方法も活用され,8-アニリノ-1-ナフタレンスルホン酸(ANS)などの蛍光プローブを用いた検出や動的光散乱を用いて会合数が増えていくようすを追跡した研究,最近では小角X線散乱をアミロイド線維形成反応の解析に適用する試みも進んでおり,線維前駆中間体の具体的な形状を提案した研究も報告されている21).これらのオリゴマー構造にはアミロイド線維形成経路上の(on-pathway)中間体の他に経路から外れた(off-pathway)ものも混在するのでその解釈は単純ではないが,アミロイド線維の核形成反応の詳細な分子機構について手がかりを与えるものである22).
2)インスリンをモデルとした核形成機構の解明
筆者らは,伝播性が発現する以前のオリゴマー様線維前駆中間体を新たに捕捉し,成熟アミロイド線維と構造比較することによって,タンパク質分子がどのような集合および構造化プロセスを経て核構造を生成し,伝播性を発現するのかについてアミノ酸残基レベルで詳細に解明することを目標とした.このために,アミロイド研究の有用なモデルタンパク質の一つであるウシ膵臓由来のインスリンに着目し,核が出現する前の中間体構造(本稿では線維前駆中間体と呼ぶことにする)を捉えその特性を解析することを試みた23).インスリンは直接疾病に関与するタンパク質ではないが,試験管内で非常に精度よくアミロイド線維を形成することから,分子レベルでのアミロイド線維形成機構を研究する上で非常に優れたモデルタンパク質である.
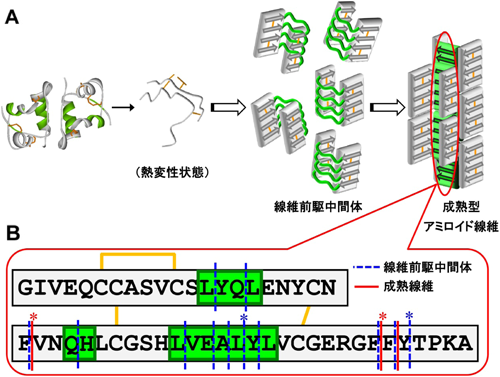
これまでの実験報告例によると,アミロイド線維形成過程で線維前駆体が顕著に蓄積するケースはまれである.そこで,主に溶媒条件に着目し線維前駆中間体を過渡的に大量蓄積するような反応系を探索した.その結果,塩濃度を高めた条件下で,試料水溶液に含まれるほぼ全量のインスリン分子が反応開始直後に微細な形状をした構造体に転換することがわかった.解析の結果,この構造体のクロスβ構造含有率は最終的なアミロイド線維構造に比べて低く,構造的に未成熟であることが確認された.さらに,アミロイド線維特有の自己触媒的な増殖性もほとんどみられないことから,この構造体はアミロイド線維核構造が形成される以前の線維前駆中間体であると結論づけた(図4A).
幸いにも,上述のアミロイド線維前駆中間体は安定性が高く寿命が長いため,詳細な構造解析を行うことができた.トリプシンによって酵素消化したときの切断箇所をもとにクロスβ構造を形成している領域としていない領域を調べ,成熟したアミロイド線維におけるクロスβ構造の形成領域と比較したところ,クロスβ構造が未完成のアミノ酸配列領域を数か所特定することができた(図4B).これらは,線維前駆中間体から成熟線維へ成熟化する際に構造形成する箇所である可能性が高く,伝播性の発現に重要な領域であると考えられる.興味深いことに,これらの領域は3節で述べたsteric zipperを作るアミノ酸配列領域と合致し24),アミロイド核構造の骨格とsteric zipper構造の構築の深い関係性を感じさせる結果であった.
以上の解析から,線維前駆中間体の捕捉および構造解析を行うことは,アミロイド線維形成初期のタンパク質の構造形成について具体的なイメージを与える有力な手段になることがわかった.過去に,寿命が長い線維前駆中間体について詳細な構造特徴を明らかにしたいくつかの興味深い解析例がある25~27).我々も実験を進め,他のアミロイド性ペプチド断片についても反応条件をうまく調節すれば線維前駆中間体を安定に観測できる可能性が徐々に明らかになってきた(未発表データ).アミロイド線維前駆中間体の捕捉は,アミロイド線維の核形成反応を明らかにするための有用な手段の一つとして活用できることが期待される.
アミロイド線維の構造研究は,ここ20年ほどで大きく展開し,プログラムされていないフォールディングとしての認識と理解も飛躍的に進んだ.アミロイド線維構造は基本的には疾病に関与しており,タンパク質分子としては異常な構造状態である.しかしながら,近年,細菌類のバイオフィルム骨格28)や哺乳類の分泌顆粒内でのペプチドホルモンの貯蔵形態29)など,正常な細胞生理機能に必要な構造として貢献する“機能性アミロイド”もいくつか発見され研究が進められている.この場合,アミロイド線維構造はプログラムされたフォールドとして有効利用されている.病気に関わるアミロイド線維と機能性アミロイドの重大な相違点は,後者においてはアミロイド線維構造が必要時に迅速に形成し,不要時には消失するというように生命システムの管理下のもとで制御されていることであろう.すなわち,アミロイド線維構造もコントロールさえかなえば決して異常な構造状態ではないのである.
本稿では,特にアミロイド線維構造を形成する初期イベントである核形成相に注目しながら話を進めたが,核形成の分子機構が明らかになればアミロイド線維の形成を未然に防ぎ人為的に制御する効果的な手立てが戦略的に開発されることが期待される.また,筆者らの進めている線維前駆中間体を経由したアミロイド線維形成反応の観察により,アミロイド線維の伝播性が発現するまでの詳細なタンパク質の集合のようすを時系列で追跡することも新たな研究展開の糸口になればと願っている.さらに最近では,アミロイド線維形成反応と一般的な物質の結晶化反応との共通性に着眼し,タンパク質が本質的に持ち合わせるアミロイド線維形成のポテンシャルを抑制し凝集を起こさない状態を物質の過飽和状態として捉えるという新しい切り口も提唱されており30,31),プログラムされていないフォールディング開始の実態がわかる日は近いかもしれない.今後の研究展開により,アミロイド線維特有の形成原理および生体ではどのようにアミロイド線維形成を抑制するのかなどについて,分子論的に明確化できるものと考えている.
謝辞Acknowledgments
ここでとりあげたインスリンの研究は,立命館大学薬学部・加藤稔教授,産業技術総合研究所・今村比呂志博士,北陸大学薬学部・山本直樹博士との共同研究であり,ここに感謝申し上げます.また,大阪大学蛋白質研究所・後藤研究室にてアミロイド線維研究を開始してから現在に至るまで多大なご意見・ご助言をいただきました後藤祐児教授に深く感謝いたします.
引用文献References
1) Dobson, C.M. (2003) Nature, 426, 884–890.
2) Sipe, J.D. & Cohen, A.S. (2000) J. Struct. Biol., 130, 88–98.
3) Eisenberg, D. & Jucker, M. (2012) Cell, 148, 1188–1203.
4) Fink, A.L. (1998) Fold. Des., 3, R9–R23.
5) Eichner, T. & Radford, S.E. (2011) Mol. Cell, 43, 8–18.
6) Berthelot, K., Cullin, C., & Lecomte, S. (2013) Biochimie, 95, 12–19.
7) Cecchi, C. & Stefani, M. (2013) Biophys. Chem., 182, 30–43.
8) Hoshi, M., Sato, M., Matsumoto, S., Noguchi, A., Yasutake, K., Yoshida, N., & Sato, K. (2003) Proc. Natl. Acad. Sci. USA, 100, 6370–6375.
9) Roychaudhuri, R., Yang, M., Hoshi, M.M., & Teplow, D.B. (2009) J. Biol. Chem., 284, 4749–4753.
10) Jahn, T.R. & Radford, S.E. (2008) Arch. Biochem. Biophys., 469, 100–117.
11) Meinhardt, J., Sachse, C., Hortschansky, P., Grigorieff, N., & Fandrich, M. (2009) J. Mol. Biol., 386, 869–877.
12) Paravastu, A.K., Leapman, R.D., Yau, W.M., & Tycko, R. (2008) Proc. Natl. Acad. Sci. USA, 105, 18349–18354.
13) Naiki, H. & Gejyo, F. (1999) Methods Enzymol., 309, 305–318.
14) Campioni, S., Monsellier, E., & Chiti, F.(2010) in Protein misfolding diseases: Current and emerging principles and therapies (Ramirez-Alvarado, M., Kelly, J.W., & Dobson, C.M. eds.), pp.3–20, John Wiley & Sons, Inc., Hoboken.
15) Hurshman Babbes, A.R., Powers, E.T., & Kelly, J.W. (2008) Biochemistry, 47, 6969–6984.
16) Murakami, K., Irie, K., Morimoto, A., Ohigashi, H., Shindo, M., Nagao, M., Shimizu, T., & Shirasawa, T. (2002) Biochem. Biophys. Res. Commun., 294, 5–10.
17) Kaji, Y., Oshika, T., Takazawa, Y., Fukayama, M., & Fujii, N. (2012) Br. J. Ophthalmol., 96, 1127–1131.
18) Goldschmidt, L., Teng, P.K., Riek, R., & Eisenberg, D. (2010) Proc. Natl. Acad. Sci. USA, 107, 3487–3492.
19) Vollers, S.S., Teplow, D.B., & Bitan, G. (2005) Methods Mol. Biol., 299, 11–18.
20) Glabe, C.G. (2008) J. Biol. Chem., 283, 29639–29643.
21) Langkilde, A.E. & Vestergaard, B. (2009) FEBS Lett., 583, 2600–2609.
22) Bemporad, F. & Chiti, F. (2012) Chem. Biol., 19, 315–327.
23) Chatani, E., Imamura, H., Yamamoto, N., & Kato, M. (2014) J. Biol. Chem., 289, 10399–10410.
24) Ivanova, M.I., Sievers, S.A., Sawaya, M.R., Wall, J.S., & Eisenberg, D. (2009) Proc. Natl. Acad. Sci. USA, 106, 18990–18995.
25) Chimon, S. & Ishii, Y. (2005) J. Am. Chem. Soc., 127, 13472–13473.
26) Laganowsky, A., Liu, C., Sawaya, M.R., Whitelegge, J.P., Park, J., Zhao, M., Pensalfini, A., Soriaga, A.B., Landau, M., Teng, P.K., Cascio, D., Glabe, C., & Eisenberg, D. (2012) Science, 335, 1228–1231.
27) Fandrich, M. (2012) J. Mol. Biol., 421, 427–440.
28) Evans, M.L. & Chapman, M.R. (2014) Biochim. Biophys. Acta, 1843, 1551–1558.
29) Maji, S.K., Perrin, M.H., Sawaya, M.R., Jessberger, S., Vadodaria, K., Rissman, R.A., Singru, P.S., Nilsson, K.P., Simon, R., Schubert, D., Eisenberg, D., Rivier, J., Sawchenko, P., Vale, W., & Riek, R. (2009) Science, 325, 328–332.
30) 後藤祐児(2013)領域融合レビュー,2, e002.
31) Ikenoue, T., Lee, Y.H., Kardos, J., Yagi, H., Ikegami, T., Naiki, H., & Goto, Y. (2014) Proc. Natl. Acad. Sci. USA, 111, 6654–6659.
著者紹介Author Profile
 茶谷 絵理(ちゃたに えり)
茶谷 絵理(ちゃたに えり)神戸大学大学院理学研究科准教授.博士(農学).
略歴1997年京都大学農学部農芸化学科卒,2002年同大学院農学研究科博士課程修了.日本学術振興会特別研究員,大阪大学蛋白質研究所特任研究員,立命館大学薬学部助教などを経て,11年4月から現職.
研究テーマと抱負アミロイド線維形成を中心に,タンパク質凝集や自己組織化の基本原理を明らかにしたい.
趣味食,園芸(始めたばかり).