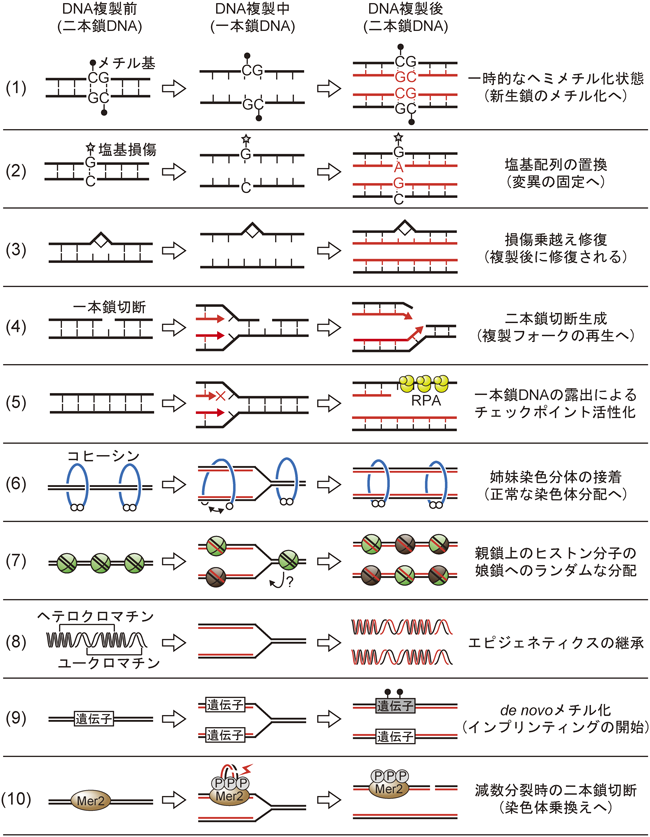
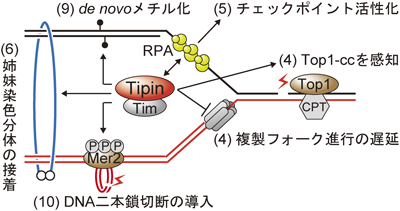
DNA複製の前後で起こる生命現象とTipinTipin solves a variety of DNA replication problems
東北薬科大学薬学部Faculty of Pharmaceutical Sciences, Tohoku Pharmaceutical University ◇ 〒981-8558 宮城県仙台市青葉区小松島四丁目4番1号4-4-1 Komatsushima, Aoba-ku, Sendai-shi, Miyagi 981-8558, Japan
発行日:2015年6月25日Published: June 25, 2015