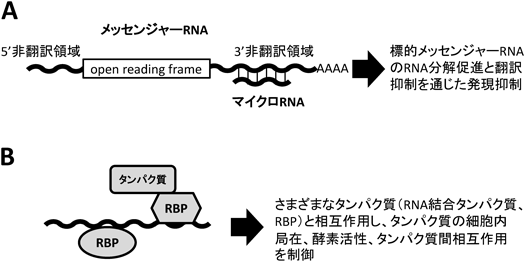
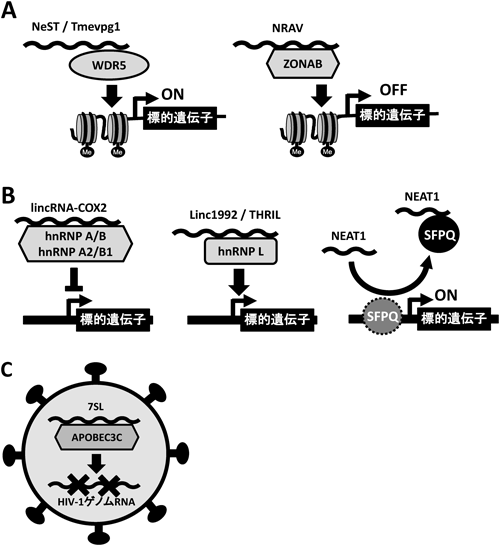
7) Carpenter, S., Aiello, D., Atianand, M.K., Ricci, E.P., Gandhi, P., Hall, L.L., Byron, M., Monks, B., Henry-Bezy, M., Lawrence, J.B., O'Neill, L.A., Moore, M.J., Caffrey, D.R., & Fitzgerald, K.A. (2013) Scinece, 341, 789–792.
8) Li, Z., Chao, T.C., Chang, K.Y., Lin, N., Patil, V.S., Shimizu, C., Head, S.R., Burns, J.C., & Rana, T.M. (2014) Proc. Natl. Acad. Sci. USA, 111, 1002–1007.
9) Gomez, J.A., Wapinski, O.L., Yang, Y.W., Bureau, J.F., Gopinath, S., Monack, D.M., Chang, H.Y., Brahic, M., & Kirkegaard, K. (2013) Cell, 152, 743–754.
10) Imamura, K., Imamachi, N., Akizuki, G., Kumakura, M., Kawaguchi, A., Nagata, K., Kato, A., Kawaguchi, Y., Sato, H., Yoneda, M., Kai, C., Yada, T., Suzuki, Y., Yamada, T., Ozawa, T., Kaneki, K., Inoue, T., Kobayashi, M., Kodama, T., Wada, Y., Sekimizu, K., & Akimitsu, N. (2014) Mol. Cell, 53, 393–406.
11) Hirose, T., Virnicchi, G., Tanigawa, A., Naganuma, T., Li, R., Kimura, H., Yokoi, T., Nakagawa, S., Bénard, M., Fox, A.H., & Pierron, G. (2014) Mol. Biol. Cell, 25, 169–183.
12) Wang, T., Tian, C., Zhang, W., Luo, K., Sarkis, P.T., Yu, L., Liu, B., Yu, Y., & Yu, X.F. (2007) J. Virol., 23, 13112–13124.
13) Winterling, C., Koch, M., Koeppel, M., Garcia-Alcalde, F., Karlas, A., & Meyer, T.F. (2014) RNA Biol., 11, 66–75.
14) Ouyang, J., Zhu, X., Chen, Y., Wei, H., Chen, Q., Chi, X., Qi, B., Zhang, L., Zhao, Y., Gao, G.F., Wang, G., & Chen, J.L. (2014) Cell Host Microbe, 16, 616–626.