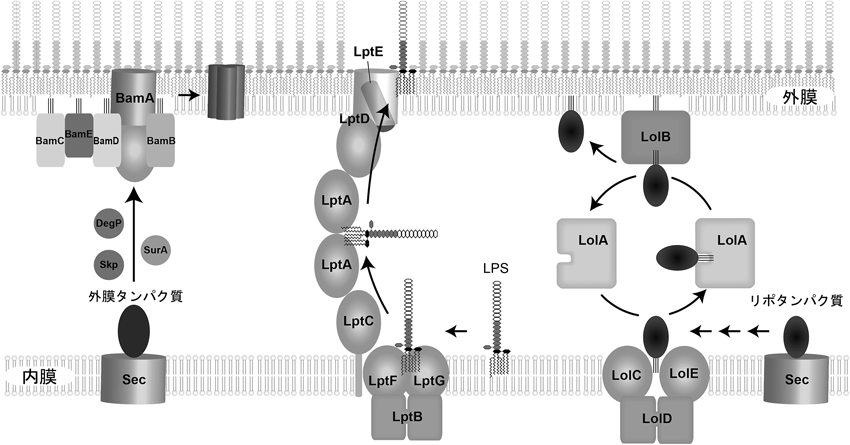
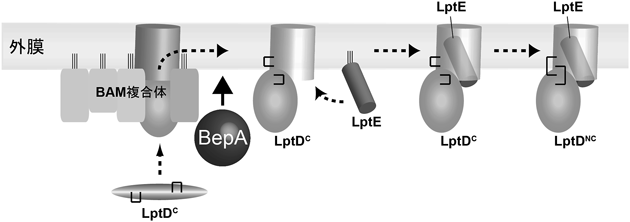
細菌外膜の生合成および維持機構Biogenesis and quality control of the outer membrane of gram-negative bacteria
盛岡大学栄養科学部Faculty of Nutritional Sciences, Morioka University ◇ 〒020-0694 岩手県滝沢市砂込808番地808 Sunakomi, Takizawa-shi, Iwate 020-0694, Japan
発行日:2015年8月25日Published: August 25, 2015