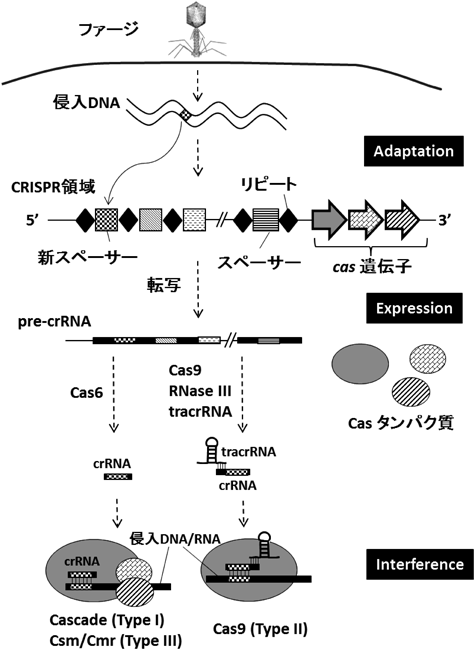
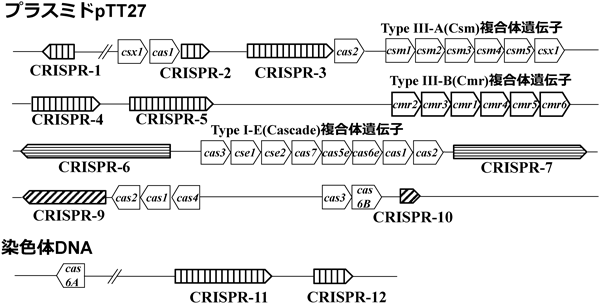
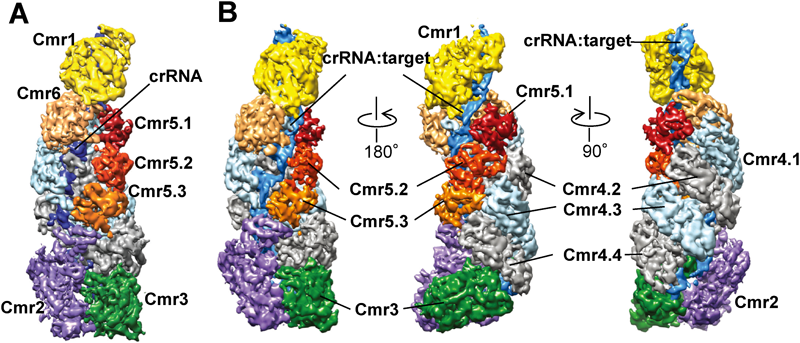
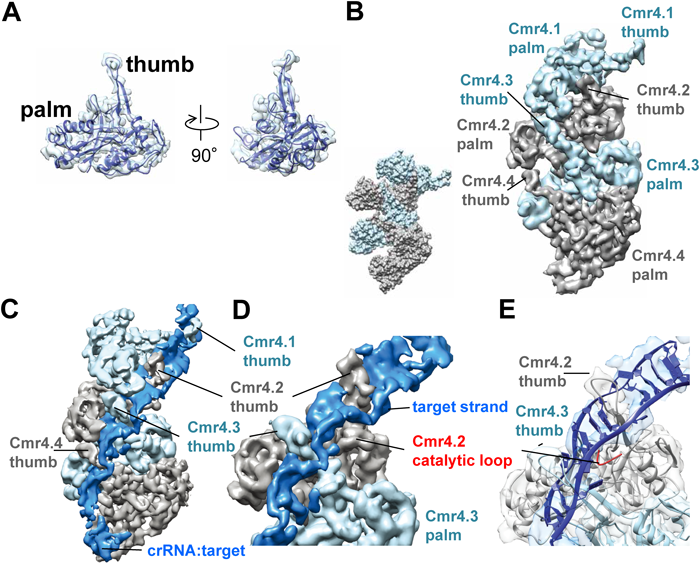
2) Makarova, K.S., Haft, D.H., Barrangou, R., Brouns, S.J., Charpentier, E., Horvath, P., Moineau, S., Mojica, F.J., Wolf, Y.I., Yakunin, A.F., van der Oost, J., & Koonin, E.V. (2011) Nat. Rev. Microbiol., 9, 467–477.
3) Agari, Y., Sakamoto, K., Tamakoshi, M., Oshima, T., Kuramitsu, S., & Shinkai, A. (2010) J. Mol. Biol., 395, 270–281.
5) Staals, R.H., Agari, Y., Maki-Yonekura, S., Zhu, Y., Taylor, D.W., van Duijn, E., Barendregt, A., Vlot, M., Koehorst, J.J., Sakamoto, K., Masuda, A., Dohmae, N., Schaap, P.J., Doudna, J.A., Heck, A.J., Yonekura, K., van der Oost, J., & Shinkai, A. (2013) Mol. Cell, 52, 135–145.
6) Taylor, D.W., Zhu, Y., Staals, R.H., Kornfield, J., Shinkai, A., van der Oost, J., Nogales, E., & Doudna, J.A. (2015) Science, 348, 581–585.
7) Benda, C., Ebert, J., Scheltema, R.A., Schiller, H.B., Baumgartner, M., Bonneau, F., Mann, M., & Conti, E. (2014) Mol. Cell, 56, 43–54.
8) Ramia, N.F., Spilman, M., Tang, L., Shao, Y., Elmore, J., Hale, C., Cocozaki, A., Bhattacharya, N., Terns, R.M., Terns, M.P., Li, H., & Stagg, S.M. (2014) Cell Reports, 9, 1610–1617.
9) Jackson, R.N., Golden, S.M., van Erp, P.B., Carter, J., Westra, E.R., Brouns, S.J., van der Oost, J., Terwilliger, T.C., Read, R.J., & Wiedenheft, B. (2014) Science, 345, 1473–1479.
11) Westra, E.R., van Erp, P.B., Kunne, T., Wong, S.P., Staals, R.H., Seegers, C.L., Bollen, S., Jore, M.M., Semenova, E., Severinov, K., de Vos, W.M., Dame, R.T., de Vries, R., Brouns, S.J., & van der Oost, J. (2012) Mol. Cell, 46, 595–605.
12) Deng, L., Garrett, R.A., Shah, S.A., Peng, X., & She, Q. (2013) Mol. Microbiol., 87, 1088–1099.
13) Peng, W., Feng, M., Feng, X., Liang, Y.X., & She, Q. (2014) Nucleic Acids Res., 43, 406–417.
14) Osawa, T., Inanaga, H., Sato, C., & Numata, T. (2015) Mol. Cell, 58, 418–430.
15) Staals, R.H., Zhu, Y., Taylor, D.W., Kornfeld, J.E., Sharma, K., Barendregt, A., Koehorst, J.J., Vlot, M., Neupane, N., Varossieau, K., Sakamoto, K., Suzuki, T., Dohmae, N., Yokoyama, S., Schaap, P.J., Urlaub, H., Heck, A.J., Nogales, E., Doudna, J.A., Shinkai, A., & van der Oost, J. (2014) Mol. Cell, 56, 518–530.