Notchシグナルは,進化的に非常によく保存された細胞の運命決定に重要なシグナル系である1,2).発生期のみならず,成体における幹細胞の増殖,分化においても重要な機能を担っていることを反映し,このシグナル系の破綻は,アラジール症候群をはじめとした遺伝性の疾患だけでなく,がんの発生および進展と転移など,多くの疾患と関連することが知られており,このシグナル系の制御機構に関する理解を深めることは,生命科学における重要な課題である3).
Notchは,I型膜貫通タンパク質受容体で,ショウジョウバエでは1種類,哺乳類では4種類の遺伝子(NOTCH1-4)が存在する2).リガンドも受容体と同様にI型膜貫通タンパク質で,ショウジョウバエでは2種類(DeltaおよびSerrate),哺乳類では5種類[Delta-like(DLL)1, 3, 4およびJagged1, 2]が同定されている.リガンドが結合するとNotchの膜貫通部位近傍の構造変化が起こる.この後,ADAMファミリーのプロテアーゼとそれに続くγ-セクレターゼによる受容体タンパク質の切断が起こり,切り出された受容体の細胞内ドメインが核内に移行し,標的遺伝子の転写を活性化する.このように,リガンドが結合後,Notchが活性化される一連の経路に関してさまざまな角度からの研究が進んでいる.しかし,リガンドの結合に直接関与するNotchの細胞外ドメインの役割についてはあまり解析が進んでいなかった.
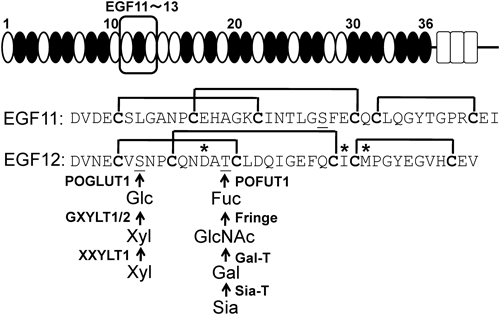
Notch細胞外ドメインは,上皮増殖因子(EGF)様リピートが数珠つなぎになっており,これが最大36回繰り返されている(図1).個々のEGFリピートには6個のシステイン残基が存在し,これらが3本のジスルフィド結合を形成することにより3次元構造が保持されている.最近の研究により,EGFリピートの糖鎖修飾がNotchの活性化制御に重要であることがわかってきた4,5).本稿では,Notch EGFリピートの糖鎖修飾によるリガンド結合の制御について,最近の2報の論文を中心に紹介し,概説したい.
まず,EGFリピートに付加することが知られているO-結合型糖鎖の構造,付加を担う糖転移酵素について述べる.
1)O-フコース(Fuc)糖鎖
Notchの機能発現に必須.O-Fuc単糖は,タンパク質O-Fuc転移酵素(POFUT1)によって,O-Fucコンセンサス配列(C2-X-X-X-X-S/T-C3,C2,C3はEGFリピート中の2番目と3番目のシステイン残基,Xはいずれかのアミノ酸を表す)中のセリンまたはトレオニン残基に,糖ヌクレオチドGDP-Fucより転移される.このO-Fuc単糖に,UDP-N-アセチルグルコサミン(UDP-GlcNAc)からGlcNAcを転移し,O-Fuc二糖(GlcNAc-Fuc)を合成する酵素としてFringeがある.Fringeは,ショウジョウバエでは1種類,哺乳類では3種類の遺伝子(Lunatic, Manic, Radical)が存在し,いずれも触媒活性を持つ.哺乳類ではさらに糖鎖が伸長され,最終的に四糖(Sia-Gal-GlcNAc-Fuc)となることもある.Fringeは,Notchシグナルの制御因子として発見され,当初は分泌タンパク質として機能解析が行われていたが,2000年になって,それがゴルジ体でNotchのO-Fuc単糖にGlcNAcを転移する糖転移酵素であること,そして,この糖鎖修飾がリガンドの結合を制御することが初めて報告された6,7).ショウジョウバエにおいて,Fringeの発現によりNotchのDelta誘導性のシグナルが増強され,一方でSerrate誘導性のシグナルが抑制されることが知られ,Fringe効果と呼ばれている8).これまで,Fringe効果の分子機構に関する明解な説明は得られていない.
2)O-グルコース(Glc)糖鎖
O-Fucと同様にNotchの機能発現に必須.O-Glc単糖は,タンパク質O-Glc転移酵素(POGLUT1/Rumi)によって,O-Glcコンセンサス配列(C1-X-S-X-P/A-C2)中のセリン残基に,糖ヌクレオチドUDP-Glcより転移される9,10).O-Glcにキシロース(Xyl)が転移し,最終的に三糖(Xyl-Xyl-Glc)となりうる.O-Glc単糖にキシロースを転移する酵素は,ショウジョウバエでは1種類(Shams),哺乳類では2種類(GXYLT1とGXYLT2)が知られ,二つ目のキシロースを転移する酵素は,哺乳類のXXYLT1が知られている.ショウジョウバエにおいては,キシロースの付加はNotchの活性化を抑制する11).
3)O-N-アセチルグルコサミン(O-GlcNAc)糖鎖
O-GlcNAc単糖は,EGFリピート特異的O-GlcNAc転移酵素(EOGT)によって,EGFリピートの5番目と6番目のシステイン残基間の特定のセリンまたはトレオニン残基に,糖ヌクレオチドUDP-GlcNAcより転移される12).この糖鎖のNotchシグナルにおける機能的重要性は不明である.
3. O-Fuc糖鎖のNotchリガンド結合における役割
我々は,リガンド結合における糖鎖の機能を解析するために,リガンド結合部位を含むヒトNOTCH1の11番目から13番目のEGFリピートを大腸菌にて発現させた13).このタンパク質hN111-13はNotchリガンドと結合することが確認されており,また,高次構造も解かれている.我々はhN111-13にin vitroで酵素的にO-FucおよびO-Glc糖鎖修飾を施した.O-Glc糖鎖については,単糖(Glc)および二糖(Xyl-Glc)を合成し,また,O-Fuc糖鎖は,単糖(Fuc),二糖(GlcNAc-Fuc),三糖(Gal-GlcNAc-Fuc),そして四糖(Sia-Gal-GlcNAc-Fuc)を必要な糖転移酵素と供与基質である糖ヌクレオチドとを混合させることによって合成した.最終的に糖鎖修飾を施したhN111-13の糖鎖の付加位置と構造を質量分析計を用いて解析した結果,O-Fuc糖鎖はEGF12のトレオニン残基T466に,O-Glc糖鎖はEGF12およびEGF13のセリン残基(S458とS496)に付加されていることを確認した(図1).
このような方法で糖鎖修飾を施したhN111-13を用い,2通りの方法でリガンド結合性を解析した.その第一は,NotchリガンドであるJagged1,DLL1あるいはDLL4を安定発現させたB16細胞またはCHO細胞に対する結合性をフローサイトメトリーにより測定する方法である.第二は,NotchリガンドのFc融合タンパク質に対するhN111-13の結合性を表面プラズモン共鳴により測定する方法である.結果を表1にまとめた.Jagged1あるいはDLL1との結合は,糖鎖修飾のないhN111-13と比較して,hN111-13のT466へのO-Fuc単糖の付加により軽度に,またGlcNAc-Fuc二糖の付加により著しく上昇したが,三糖または四糖への糖鎖伸長は結合に影響を与えなかった.一方,DLL4との結合に対してはO-Fuc糖鎖の影響は検出されなかった.増強効果が認められない原因としては,糖鎖修飾のないhN111-13へのDLL4の結合がもともと高いためであると考えられた.また,hN111-13へのO-Glc糖鎖の付加は,Jagged1,DLL1あるいはDLL4への結合性に影響しなかった.
表1 O-Fuc糖鎖付加のNotchリガンド結合への影響13)| 糖鎖付加 | Notchリガンド |
|---|
| Jagged1 | DLL1 | DLL4 |
|---|
| Fuc | ↑ | ↑ | — |
| GlcNAc-Fuc | ↑↑ | ↑↑ | — |
| Gal-GlcNAc-Fuc | — | — | — |
| Sia-Gal-GlcNAc-Fuc | — | — | — |
| ↑:やや増強,↑↑:著しく増強,一影響なし. |
O-Fuc糖鎖の付加により,Notchリガンドの結合性が変化したので,次にT466にO-Fuc単糖,あるいは二糖を付加させたhN111-13のX線結晶構造解析を行い,糖鎖修飾のないhN111-13と構造を比較した13).その結果,2点の興味深い事実が明らかとなった.O-Fuc単糖は,リガンド結合に関与するEGF12のβ-シートから突き出すように位置し,T466の側鎖のコンホメーションは“m”ロータマー(回転異性体)で,構造的に安定に存在していた.このO-Fuc糖鎖による構造の安定化は,O-FucのC6メチル基と,EGF12中のI477およびM479との相互作用によることが示唆された.GlcNAc-Fuc二糖の場合には,GlcNAcはリガンド結合表面を2倍近くに拡大させ,EGF12中のD464およびM479と相互作用することが明らかになった.EGFリピート中のO-Fuc糖鎖の構造的安定性に関する知見は,Notchシグナルの制御における糖鎖付加の意義にとどまらず,より広い意味で,タンパク質と糖鎖の相互作用,そして,糖鎖が特定の分子内存在様式を持つことによってタンパク質の機能を制御する例として重要である.二つ目の重要な点は,個々のEGFリピートのフォールディング状態は3本のジスルフィド結合により,そのフォールディング状態が“かっちりと”極(き)められているため想定されていたことであるが,O-Fuc糖鎖の付加は,T466を含むEGF12の構造,EGFリピート間のつなぎ目の構造,およびhN111-13全体の構造を変化させないことが実証されたことである.
以上より,Notch1 EGF12のT466に付加されたO-Fuc糖鎖は,リガンド結合表面を拡大し,リガンドとの相互作用に直接的に関与することが強く示唆されたが,このことを証明する最も有効な手段は,Notchとそのリガンドとの共結晶化による相互作用の解析であると考えられた.
4. DLL4とNotch1との共結晶化によって示されたNotchリガンド結合制御の分子基盤
ごく最近,興味深い実験結果が,構造生物学を専門とするスタンフォード大のグループから報告された.彼らは,DLL4とN111-13との複合体の共結晶化に成功したのである14).以下,その実験手法と結果を概説したい.
一般に,共結晶化には分子複合体が安定,つまり分子間の相互作用が強い方が有利である.裏を返せば,Notch-リガンド間のそれほど強くない相互作用が,これまで共結晶化を阻んできた一因である.彼らは,この問題を解決するために,アフィニティー・マチュレーションにより高親和性のNotchリガンドDLL4を作出した.具体的には,DLL4のNotchへの結合に必要と想定されている部分(N末端のMNNLドメイン,DSLドメイン,そして2番目のEGFリピートまで)を含むDLL4(N-EGF2)および,さらに三つのEGFリピートを含むDLL4(N-EGF5)と酵母細胞表面タンパク質Aga2との融合タンパク質を鋳型として,DLL4(N-EGF5)の変異体ライブラリーを作製し,Notch11-14への結合性を指標としたバイオパニングを複数回繰り返した.得られたDLL4変異体を昆虫細胞で発現,精製し,Notch1に対する親和性を表面プラズモン共鳴法によって解析した.野生型のDLL4(N-EGF2)は,N11-14,N110-14,N111-13に対して,それぞれ12.7 µM,8.6 µM,7.5 µMの解離定数(Kd)を示したのに対し,SLP変異(G28S,F107L,およびL206P)を含む第一世代変異体DLL4SLP(N-EGF2)は,それぞれ444 nM,539 nM,262 nMのKdを,また,SLP変異に加えて四つの変異(N118I, I143F, H194Y, K215E)を含む第二世代変異体DLL4E12(N-EGF2)は,それぞれ56.3 nM,67.8 nM,59.7 nMのKdを示した.高親和性DLL4変異体がNotchシグナルを誘導する機能を失っていないことは,培養細胞を用いたリポーターアッセイによって確認され,DLL4SLP(N-EGF2)は,野生型と同程度のNotchシグナルの活性化を誘導し,DLL4E12(N-EGF2)は野生型に比べて5倍程度,Notchシグナルの活性化を誘導した.
彼らは,高親和性変異体DLL4SLP(N-EGF1)あるいはDLL4SLP(N-EGF2)と昆虫細胞で発現させたN111-13との共結晶化およびX線構造解析に成功した(解像度は,それぞれ2.3 Åおよび3.4 Å).DLL4SLP(N-EGF2)とN111-13との複合体において,両者は逆平行におよそ1100 Å2の面積を共有して結合し,ここにN111-13のEGF11およびEGF12のアミノ酸残基,そしてT466に付加したO-Fuc糖鎖も存在していた.Notchとリガンドとの結合はカルシウムに依存することが知られていたが,カルシウムは結合に直接関与しておらず,NotchのEGFリピート間に存在するカルシウム結合配列に結合していたことから,カルシウムはN111-13の構造を安定化させることによってリガンド結合に寄与していることがわかった.また,共結晶中のN111-13およびリガンドに結合していないN111-13の構造を比較すると,その差は非常に小さく,DLL4との結合に伴ってN111-13は有意な構造変化を起こしていないことが示唆された.この複合体において,DLL4のMNNLドメインとN111-13のEGF12(Site 1)の間,そしてDLL4のDSLドメインとN111-13のEGF11(Site 2)の間に相互作用があることが明らかになった.2か所で相互作用することは,何を意味しているのであろうか? 彼らは,Notchおよびリガンドのアミノ酸配列を比較し,保存性の高いSite 2の相互作用は,互いにNotchとそのリガンドであることを確認するのに役立ち,より多様なアミノ酸配列の組み合わせからなるSite 1を介する相互作用によって特異性が規定され,O-Fuc糖鎖がその親和性を制御するために重要な役割を担っているのではないかと推測している.
Site 1において,O-Fuc単糖は結合表面の約80%を占め,糖鎖–タンパク質間相互作用ネットワークの中心に位置していた.DLL4のMNNLドメインのH64とY65の側鎖芳香環がN111-13のEGF12中のO-Fuc単糖と相互作用し,O-Fuc単糖の水酸基がMNNLドメインのY65の主鎖のカルボニル基と水素結合を形成していた.共結晶化されたN111-13のEGF12中のO-Fuc糖鎖は単糖であったため,彼らはFringeによってGlcNAcがO-Fucに付加された二糖構造をモデリングにより再構築し,DLL4との結合に与える影響を考察した.その結果,O-Fuc二糖は,我々が実験的に明らかにした構造と同様に,N111-13のM479およびD464と相互作用することが示唆された.また,DLL4のY65と相互作用することも示唆された.このように,O-Fuc糖鎖がNotch1とDLL4との相互作用部位に介在していることが示されたが,この糖鎖の存否,そしてO-Fuc四糖構造までも含めた糖鎖の伸長度の違いが,親和性にどの程度影響を及ぼすかについては今後の課題である.我々の実験では,前に述べたようにDLL4は糖鎖修飾のないN111-13に対する結合性が高く,糖鎖付加による変化は観察されなかった.
興味深いことに,彼らはN111-13のEGF11のS435がO-Glcで修飾されており,これがSite 2においてDLL4のD218とQ219と相互作用していることを見いだした.これまでNotch1のEGF11における糖鎖修飾は知られていなかった.S435はEGF11中の3番目と4番目のシステイン残基の間に存在し,既知のO-Glcコンセンサス配列には当てはまらず,未知のO-Glc糖転移酵素が存在すると考えられる.コンセンサス配列内のO-Glc糖鎖はキシロースによって伸長される.S435のO-Glcも同様に伸長されるとすれば,彼らが指摘しているとおり,Notchリガンド相互作用の新たな制御メカニズムの発見につながるかもしれない.
糖鎖の生物学的機能を追求するための生化学的研究が,複雑かつ巧妙なNotchシグナル系の制御メカニズムの一端を明らかにしつつある5).これまで述べてきたように,Notch1のリガンド結合部位に存在するO-Fuc糖鎖は,EGF12のポリペプチドと相互作用することによって安定したリガンド結合表面を形成し,Jagged1およびDLL1との結合親和性の上昇に寄与している13).さらに,Notch1とDLL4との共結晶構造解析により,O-Fuc単糖がNotch1のリガンド結合表面に存在し,リガンドとの結合に直接関与していることが確かめられた14).
哺乳類には,4種類のNotchが存在し,これらの一次構造はよく保存されているが,細胞外部位のEGFリピートの繰り返しの回数が異なっており,このことの意味は理解されていない.今回概説した研究結果は,いずれもNotchリガンド結合部位に限局したものであり,この部位以外のEGFリピートが受容体活性化において何らかの役割を担っている可能性もある.実際,Notch1の活性化にEGF26とEGF27に付加されたO-Fuc糖鎖が必要であることが示唆されているので15),Notch細胞外領域全体からその分子機能を捉えることが次の課題であろう.
謝辞Acknowledgments
ここに紹介した研究の一部は,ニューヨーク州立ストーニーブルック大学のBob Haltiwanger博士の研究室で行われたものです.私の研究活動を支えて下さったHaltiwanger博士,研究室員の皆様,そして私の家族に深く感謝致します.
引用文献References
1) Artavanis-Tsakonas, S., Rand, M.D., & Lake, R.J. (1999) Science, 284, 770–776.
2) Kopan, R. & Ilagan, M.X. (2009) Cell, 137, 216–233.
3) Ntziachristos, P., Lim, J.S., Sage, J., & Aifantis, I. (2014) Cancer Cell, 25, 318–334.
4) Takeuchi, H. & Haltiwanger, R.S. (2010) Semin. Cell Dev. Biol., 21, 638–645.
5) Takeuchi, H. & Haltiwanger, R.S. (2014) Biochem. Biophys. Res. Commun., 453, 235–242.
6) Bruckner, K., Perez, L., Clausen, H., & Cohen, S. (2000) Nature, 406, 411–415.
7) Moloney, D.J., Panin, V.M., Johnston, S.H., Chen, J., Shao, L., Wilson, R., Wang, Y., Stanley, P., Irvine, K.D., Haltiwanger, R.S., & Vogt, T.F. (2000) Nature, 406, 369–375.
8) Rana, N.A. & Haltiwanger, R.S. (2011) Curr. Opin. Struct. Biol., 21, 583–589.
9) Acar, M., Jafar-Nejad, H., Takeuchi, H., Rajan, A., Ibrani, D., Rana, N.A., Pan, H., Haltiwanger, R.S., & Bellen, H.J. (2008) Cell, 132, 247–258.
10) Takeuchi, H., Fernandez-Valdivia, R.C., Caswell, D.S., Nita-Lazar, A., Rana, N.A., Garner, T.P., Weldeghiorghis, T.K., Macnaughtan, M.A., Jafar-Nejad, H., & Haltiwanger, R.S. (2011) Proc. Natl. Acad. Sci. USA, 108, 16600–16605.
11) Lee, T.V., Sethi, M.K., Leonardi, J., Rana, N.A., Buettner, F.F., Haltiwanger, R.S., Bakker, H., & Jafar-Nejad, H. (2013) PLoS Genet., 9, e1003547.
12) Sakaidani, Y., Nomura, T., Matsuura, A., Ito, M., Suzuki, E., Murakami, K., Nadano, D., Matsuda, T., Furukawa, K., & Okajima, T. (2011) Nat. Commun., 2, 583.
13) Taylor, P., Takeuchi, H., Sheppard, D., Chillakuri, C., Lea, S.M., Haltiwanger, R.S., & Handford, P.A. (2014) Proc. Natl. Acad. Sci. USA, 111, 7290–7295.
14) Luca, V.C., Jude, K.M., Pierce, N.W., Nachury, M.V., Fischer, S., & Garcia, K.C. (2015) Science, 347, 847–853.
15) Rampal, R., Arboleda-Velasquez, J.F., Nita-Lazar, A., Kosik, K.S., & Haltiwanger, R.S. (2005) J. Biol. Chem., 280, 32133–32140.
著者紹介Author Profile
 竹内 英之(たけうち ひでゆき)
竹内 英之(たけうち ひでゆき)ニューヨーク州立大学ストーニーブルック大学生化学細胞生物学Research Assistant Professor. 博士(薬学).
略歴1972年埼玉県に生まれる.96年東京大学薬学部薬学科卒業.2001年東京大学大学院薬学系研究科機能薬学専攻修了.東京大学大学院薬学系研究科では,入村達郎教授に師事.06年よりHaltiwanger博士に師事.08年より現職.
研究テーマと抱負O-結合型糖鎖の生物学的重要性の解明.現在は,比較的単純な構造からなるO-Glc糖鎖が,Notchシグナルを制御する仕組みを明らかにすることに注力している.2015年夏より,ジョージア大学Complex Carbohydrate Research Centerに異動予定.
趣味スポーツ,特に野球とテニス.