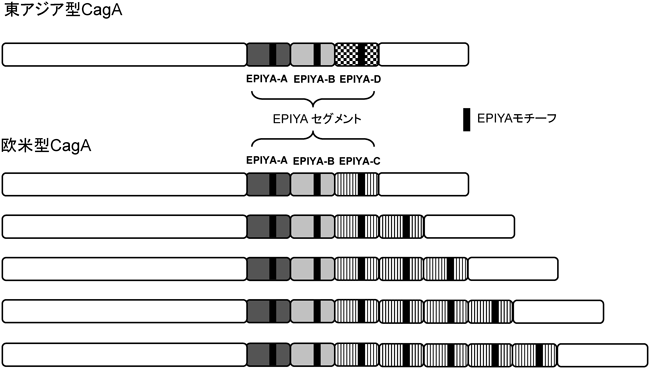
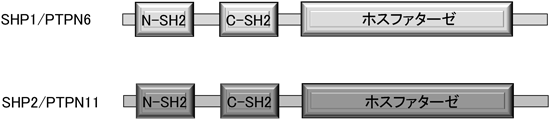
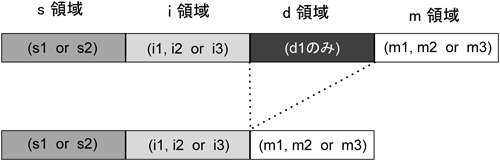
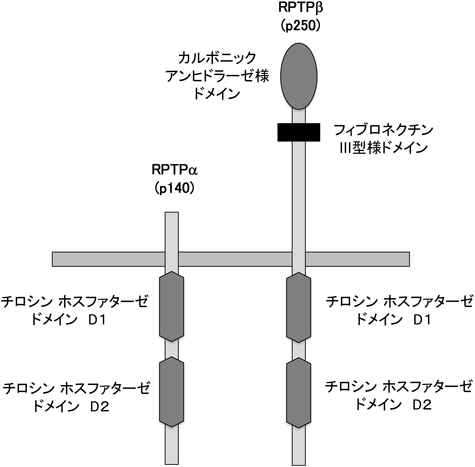
ピロリ菌発がんとチロシンホスファターゼHelicobacter pylori-mediated carcinogenesis and tyrosine phosphatases
東京大学大学院医学系研究科Graduate School of Medicine, The University of Tokyo ◇ 〒113-0033 東京都文京区本郷七丁目3番1号Hongo 7-3-1, Bunkyo-ku, Tokyo 113-0033, Japan
発行日:2015年10月25日Published: October 25, 2015