4) Schulz, M.J., Tecedor, L., Chang, M., & Davidson, B.L. (2011) Trends Neurosci., 34, 401–410.
5) Suzuki, Y., Nanba, E., Matsuda, J., Higaki, K., & Oshima, A. (2008) The Online Metabolic and Molecular Bases of Inherited Disease (Valle, D., Beaudet A.L., Vogelstein, B., Kinzler, K.W., Antonarakis, S.F., & Ballabio, A., eds.), pp. 1–101, McGraw-Hill, New York.
6) Okumiya, T., Ishii, S., Takenaka, T., Kase, R., Kamei, S., Sakuraba, H., & Suzuki, Y. (1995) Biochem. Biophys. Res. Commun., 214, 1219–1224.
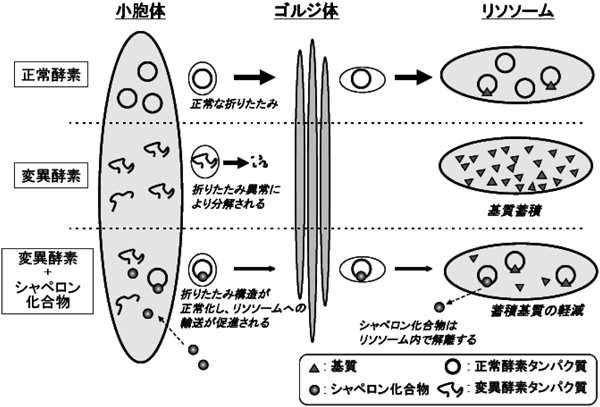
7) Fan, J.Q., Ishii, S., Asano, N., & Suzuki, Y. (1999) Nat. Med., 5, 112–115.
8) Giugliani, R., Waldek, S., Germain, D.P., Nicholls, K., Bichet, D.G., Simosky, J.K., Bragat, A.C., Castelli, J.P., Benjamin, E.R., & Boudes, P.F. (2013) Mol. Genet. Metab., 109, 86–92.
9) Matsuda, J., Suzuki, O., Ohshima, A., Yamamoto, Y., Noguchi, A., Takimoto, K., Itoh, M., Yasuda, Y., Ogawa, S., Sakata, Y., Nanba, E., Higaki, K., Ogawa, T., Tominaga, L., Ohno, K., Iwasaki, H., Watanabe, H., Brady, R.O., & Suzuki, Y. (2003) Proc. Natl. Acad. Sci. USA, 100, 15912–15917.
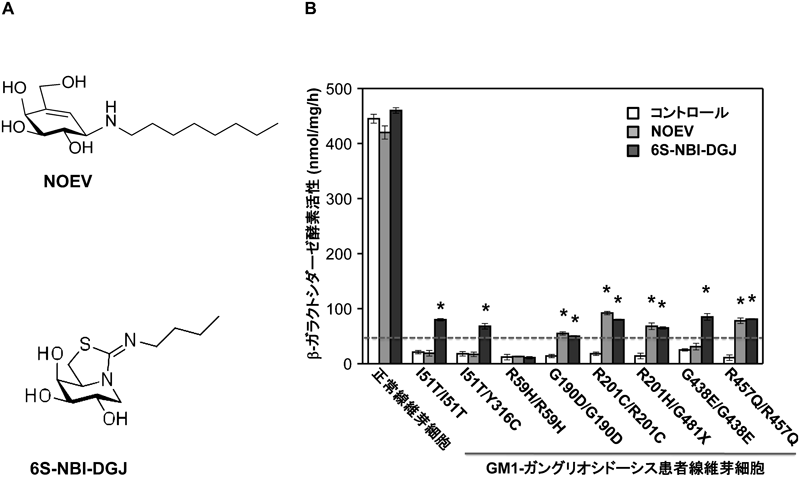
10) Takai, T., Higaki, K., Aguilar-Moncayo, M., Mena-Barragán, T., Hirano, Y., Yura, K., Yu, L., Ninomiya, H., García-Moreno, I., Sakakibara, Y., Ohno, K., Nanba, E., Ortiz-Mellet, C., García-Fernández, J.M., & Suzuki, Y. (2013) Mol. Ther., 21, 526–532.
11) Suzuki, H., Ohto, U., Higaki, K., Mena-Barragan, T., Aguilar-Moncayo, M., Ortiz-Mellet, C., Nanba, E., Garcia-Fernandez, J.M., Suzuki, Y., & Shimizu, T. (2014) J. Biol. Chem., 289, 14560–14568.
12) Iwasaki, H., Watanabe, H., Iida, M., Ogawa, S., Tabe, M., Higaki, K., Nanba, E., & Suzuki, Y. (2006) Brain Dev., 28, 482–486.
13) Higaki, K., Li, L., Bahrudin, U., Okuzawa, S., Takamura, A., Yamamoto, K., Adachi, K., Paraguison, R.C., Takai, T., Ikehata, H., Tominaga, L., Hisatome, I., Iida, M., Ogawa, S., Matsuda, J., Ninomiya, H., Sakakibara, Y., Ohno, K., Suzuki, Y., & Nanba, E. (2011) Hum. Mutat., 32, 843–852.
14) Suzuki, Y., Ichinomiya, S., Kurosawa, M., Ohkubi, H., Watanabe, H., Iwasaki, H., Matsuda, J., Noguchi, Y., Takimoto, K., Itoh, M., Tabe, M., Iida, M., Kubo, T., Ogawa, S., Nanba, E., Higaki, K., Ohno, K., & Brady, R.O. (2007) Ann. Neurol., 62, 671–675.
15) Suzuki, Y., Ichinomiya, S., Kurosawa, M., Matsuda, J., Ogawa, S., Iida, M., Kubo, T., Tabe, M., Itoh, M., Higaki, K., Nanba, E., & Ohno, K. (2012) Mol. Genet. Metab., 106, 92–98.
16) Porto, C., Ferrara, M.C., Meli, M., Acampora, E., Avolio, V., Rosa, M., Cobucci-Ponzano, B., Colombo, G., Moracci, M., Andria, G., & Parenti, G. (2012) Mol. Ther., 20, 2201–2211.