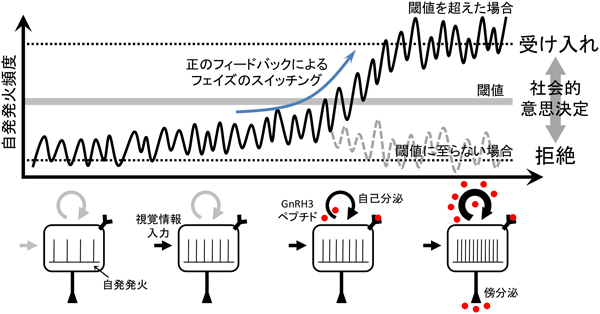
「社会脳」の進化的起源の解明を目指して―社会的意思決定を担う終神経GnRH3ニューロンExploring evolutionary roots of “social brain”: Central role of terminal nerve GnRH3 neurons in social decision making
1 RIKEN-MIT Center for Neural Circuit Genetics (Tonegawa Lab), Picower Institute for Learning and Memory, Massachusetts Institute of Technology ◇ 43 Vassar Street, Bldg. 46-6461, Cambridge, MA 02139, United States of America
2 岡山大学大学院自然科学研究科Graduate School of Natural Science and Technology, Okayama University ◇ 〒700-8530 岡山市北区津島中三丁目1番1号Tsushimanaka 3-1-1, Kita-ku, Okayama-shi, Okayama 700-8530, Japan
発行日:2015年10月25日Published: October 25, 2015