多細胞生物の同一個体内においてほぼすべての細胞は同じゲノムを有しているのにも関わらず,それぞれの細胞は発生時期やその属する臓器・組織に特有の異なる遺伝子発現プロファイルを示している.このような遺伝子発現の時空間的な制御(時期特異的制御,組織特異的制御)は多細胞生物の発生,臓器形成等の生命現象において中心的な役割を果たしており,そしてその制御は主に転写レベルで行われていると考えられている1,2).したがって,時空間的な転写制御の分子メカニズムの詳細を理解することは分子生物学の大きな課題の一つである.
組織特異的,発生段階特異的な遺伝子発現プロファイルはエンハンサーDNA配列やそれを認識する特異的なDNA結合性転写因子群によって規定されていると考えられてきた.しかしながら,近年の研究からこれらに加えてTFIID等の基本転写因子群の複合体構成因子の組み合わせの多様性が時空間特異的な転写制御に寄与していることが徐々に明らかとなってきた.さらに,このような組織特異的な複合体の組み換えモデルはクロマチンリモデリング因子複合体やメディエーター複合体といった転写アクティベーターにまで拡大しつつある.最近筆者らは神経細胞特異的なヒストン脱メチル化酵素複合体を同定し,ヒストン修飾因子複合体も組織特異的な組み合わせをとりうること,またこの複合体が組織特異的な転写制御に寄与することを報告した.最近の組織特異的転写制御研究の知見とともにこれを紹介したい.
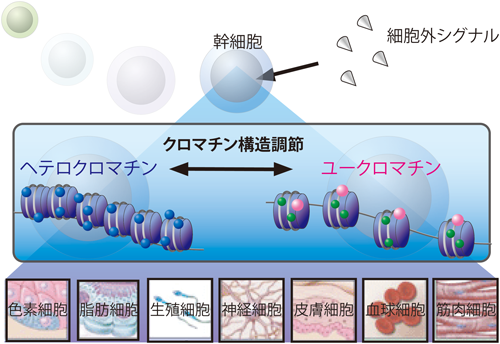
多細胞生物の時空間的転写制御を支える分子基盤としてDNAを取り巻くクロマチン環境とクロマチン構造変換の制御があげられる.多細胞生物をはじめとする真核細胞のDNAはトータル1.7 mもの長さのDNAがヒストンタンパク質に巻きついたヌクレオソームを形成しクロマチン構造をとることにより高度に折りたたまれ,直径約5 µmほどの細胞核内に収納されている3).そのため,細胞内のDNAは高度に折りたたまれ凝縮された構造をとっており転写が不活性化している状態にあると考えられる.実際,ヌクレオソームを形成したDNAは試験管内反応においても転写反応を強く阻害することが古くから知られている4).そのため,真核細胞の転写制御においてはこのクロマチン構造を適切に調節するメカニズムが存在することになる.
クロマチン構造は細胞核内において一様ではなく,ヌクレオソーム間隔が粗で弛緩している状態のユークロマチンと,ヌクレオソームが密になりDNAが凝縮した状態のヘテロクロマチンの2種類に大別される.ユークロマチン領域では転写因子がDNAにアクセスすることが可能であり転写が活発に行われている.実際,活性化されたプロモーターのDNAはヌクレアーゼによる切断を受けやすいという事実から,ヌクレオソームの密度と転写活性化には密接な関連があると考えられてきた5).これに対し,ヘテロクロマチン領域ではDNAが密に凝縮されているため転写因子がDNAに結合できず転写が不活性になっていると考えられている.
この核内のクロマチン構造の粗密のパターンは,多細胞生物のそれぞれの特化した組織および細胞において特異的なパターンが構築されており,これが組織特異的な転写制御において重要な役割を担っていることがわかる.ではそれぞれの細胞においてどのように特異的なクロマチン構造調節が実現されているのであろうか.それは個々の細胞において“エピゲノム”と称されるクロマチンの状態が異なるからに他ならない6)(図1).
エピゲノムとは後天的に修飾されたゲノムのことであり,つまりクロマチンの構造や修飾状態の総称である.このエピゲノム調節には現在までに大きく分けて,(1)DNAの化学修飾(メチル化,ヒドロキシメチル化等),(2)ヒストンタンパク質の翻訳後修飾(アセチル化,メチル化,リン酸化,ユビキチン化等),(3)ATP依存性のクロマチンリモデリング,(4)ヒストンシャペロンによるヒストンタンパク質の置き換え,(5)non-coding RNAによる制御,の5種類の制御機構が知られている.
それぞれの機構は固有の酵素活性を持った転写共役因子と呼ばれる一連の因子群により制御されている.そしてこれら転写共役因子群はこれまでの研究から,多くの場合さまざまな因子と核内で複合体を形成することにより機能していることがわかってきた7,8).これまでクロマチンリモデリング因子BRG1(Brahma-related gene 1)やヒストンメチル化酵素MLL1(mixed lineage leukemia 1)をはじめ多くの転写共役因子複合体の構成因子が生化学的に単離・同定されてきた.しかしながら,一般的に複合体の精製・同定には多量の核抽出液(総タンパク質量100 mg~)を出発材料として要するため,多くの場合これら複合体はヒト由来HeLa細胞やHEK293細胞,もしくはショウジョウバエ由来S2細胞といった大量培養や核抽出液の調製が比較的しやすい細胞株が精製材料として用いられてきた経緯がある.そのため,実際に個々の特異的な細胞においてもこれら転写共役因子が同様の構成因子と複合体を形成し存在しているのかについては不明であった.
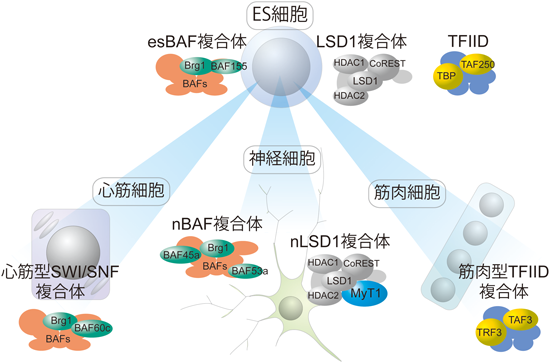
しかしながら,近年複数のグループから転写に関連する複合体が組織によってその構成因子を変える例が報告されてきている.まず一つ目の例は,筋分化の際に基本転写因子TFIID複合体のコアコンポーネントであるTBP(TAT A-binding protein)が消失することである9).これまで組織特異的TAF(TBP-associated factor)の報告はあったがTBPが筋分化依存的に消失するということはTFIID複合体が細胞分化依存的に動的に再編成されていることを意味する.実際,筋分化後の細胞ではTBPに代わってTBPによく似たTRF3という因子が代替的に機能していることが明らかとなった.
また,別のグループからの報告では,ATPアーゼBRG1をコアコンポーネントとするSWI/SNF型のクロマチンリモデリング複合体の構成因子(BAF: BRG1-associated factor)が細胞分化に伴って複合体構成因子の再編成を行うことが示されている10).たとえば,BAF60cサブユニットを含んだBAF複合体は心筋細胞に特異的に存在し心筋細胞の成熟化に必要なことが示唆されている.また神経前駆細胞からニューロンへの分化の際に,BAF45aやBAF53aサブユニットが別のサブユニットへと交換されていることが報告されていて,トランスジェニックマウスを使った実験からこのサブユニット交換は神経分化に必須であることが証明されている.さらに,ES細胞特異的なesBAF複合体はES細胞の未分化維持や細胞増殖に必要で,そのノックダウンは自発的なES細胞分化を生じさせてしまう.このようにATP依存性クロマチンリモデリング因子複合体が組織ごとに特異的な複合体を形成し,さらにそのことが正常な細胞分化にとって必須の過程であることがわかってきた.しかしながら,これらの事象がヒストン修飾因子複合体にも起こりうることなのかという点についてはこれまで報告がなされていなかった.
そこで我々は,特異的細胞のモデルとして神経細胞を用いてヒストン修飾因子LSD1(lysine specific demethylase 1)複合体の精製・同定を試みた.LSD1はヒストンに対して直接脱メチル化反応を行う酵素として2004年に初めて同定された因子であり神経細胞をはじめ広範な細胞において発現が認められる11).LSD1の形成する複合体はHEK293細胞より精製されており,CoREST,HDAC1/2を含む複数のサブユニットが同定されている12).我々は神経芽腫由来細胞株Neuro2a細胞にてFLAG-LSD1を安定的に発現する細胞を樹立した後,この細胞の大量培養,核抽出液の調製およびFLAG-LSD1複合体の精製を行った.アフィニティーカラム精製,イオン交換カラム精製,グリセロール密度勾配遠心によるサイズ分画の3段階精製によりNeuro2a細胞におけるLSD1複合体の単離に成功した.さらに,精製されたLSD1複合体分画を質量分析計(LC-MS/MS)にて解析することにより,その複合体構成因子の同定を行った.得られた構成因子を過去のLSD1複合体の報告と比較検討した結果,このNeuro2a細胞に特異的なLSD1複合体構成因子の一つとしてMyT1(myelin transcription factor 1)を見いだした13).
MyT1はミエリン鞘で豊富に発現するPLP1(proteolipid protein 1)遺伝子のプロモーター領域に結合する因子としてヒト胎児脳のcDNAライブラリーからクローニングされたC2HC型のジンクフィンガー因子である14).しかしながら,転写調節能を含めたその詳細な分子機能はこれまでほとんど不明であった.今回の我々のNeuro2a細胞を用いた実験の結果から,MyT1はLSD1と直接結合することでCoRESTやHDAC1/2と生化学的に安定な複合体を形成することが明らかとなった.さらにマウス脳を用いた組織切片の抗体染色を行ったところ,ニューロンマーカーTuJ1陽性細胞の細胞核においてMyT1およびLSD1の共局在が確認されたことから,培養細胞のみならず生体内でもこのような複合体が存在していることが強く示唆された.そこで我々はMyT1とLSD1が形成するこの神経細胞特異的な複合体をnLSD1複合体(neural cell-specific LSD1 complex)と命名しさらなる解析を進めた.
nLSD1複合体が制御する遺伝子群を明らかにするために,siRNAとマイクロアレイを組み合わせた標的遺伝子探索を行った.その結果,nLSD1複合体はPTEN遺伝子をはじめとする細胞増殖関連遺伝子の他,St18(Myt3)等の神経細胞における機能が報告されている遺伝子群の発現を負に制御していることが判明した.実際に,MyT1およびLSD1のノックダウンによりPTEN遺伝子の発現は上昇し細胞増殖の抑制が認められた.さらに,クロマチン免疫沈降法によりPTEN遺伝子プロモーター上にnLSD1複合体が存在し活性化クロマチンの指標の一つであるヒストンH3K4me2を脱メチル化していることも確認された.
以上の結果から,少なくとも神経細胞には特異的なLSD1複合体が存在し,神経細胞特異的な転写制御を行っていると考えることができる.したがって,組織特異的な転写関連複合体が組織特異的転写制御に寄与しているという概念は,これまでの基本転写因子複合体,クロマチンリモデリング複合体に加えてヒストン修飾因子複合体にも適用できるようである.このことを一般化すると,各種転写関連複合体において,組織ごとに異なる複合体構成因子が存在し,それぞれの組織で特有の複合体を形成し組織特異的な転写制御を可能にしていると推測することができる(図2).
転写共役因子複合体の構成因子がなぜ組織特異的に変換されるかの理由についてはまだ明らかではない.一つの可能性として組織特異的な構成因子が複合体とDNA結合性転写因子との相互作用のアダプターとなっていることがあげられる.つまり,組織特異的なアダプターを持つことで同じ活性を持った複合体が,組織ごとに異なる転写因子と結合することが可能となり特異的な標的遺伝子プロモーター上にリクルートされる.この分子機構によって,組織特異的なクロマチン構造調節を説明することができる.このアイデアによるとMyT1もこのアダプターの一種と考えられる.この仮説を支持するデータとして我々はMyT1をノックダウンするとLSD1のPTEN遺伝子プロモーターへのリクルートが減弱するという結果も得ている.
iPS細胞をはじめとするダイレクトリプログラミングの発見から,それぞれの細胞に固有のマスター転写因子が特異的な遺伝子発現パターンを獲得するのにきわめて重要なことは明らかである.しかしながら,これらのリプログラミング時においても転写共役因子群によりエピゲノム調節が行われていると考えられるが,はたしてどのような複合体構成因子をとっているのかはまったく未知である.これまでの知見から,リプログラミング時にも特異的な複合体構成因子が厳密に使い分けられることによってそれぞれ目的の細胞に分化・脱分化していることが予想される.
興味深いことに,皮膚線維芽細胞から機能的なニューロンへリプログラミングされる際に必要な三つの因子のうちの一つがMyT1のサブタイプであるMyT1Lであることが近年明らかになっている15).筆者らの実験から,MyT1LもMyT1と同様にLSD1と複合体形成することが判明している.したがって,線維芽細胞からニューロンへのリプログラミング時にもnLSD1複合体による標的遺伝子のヒストン脱メチル化とそれに伴う転写調節が重要な役割を果たしていることが大いに予想されるところである.
また,ヒストン修飾因子はがんをはじめ多くの疾患においてその発症に関与していると考えられ創薬標的として提案されている.たとえば,LSD1は前立腺がんや急性骨髄性白血病(AML),うつ病との関連が指摘されている.しかしながら,LSD1をはじめとするヒストン修飾因子複合体の活性サブユニットは多くの場合広範な組織において発現しており,これらをターゲットとした薬剤は重篤な副作用も予想される.この点において,これらエピゲノム調節因子が複合体を形成していること,そしてその複合体は組織によって特異的な構成因子をとることを考慮すると,MyT1のような組織特異的なサブユニットを創薬ターゲットとすることでより特異的に標的組織において効果を発揮する薬剤の開発につながる可能性がある.
引用文献References
1) Mitchell, P.J. & Tjian, R. (1989) Science, 245, 371–378.
2) Roeder, R.G. (2003) Nat. Med., 9, 1239–1244.
3) Campos, E.I. & Reinberg, D. (2009) Annu. Rev. Genet., 43, 559–599.
4) Laybourn, P.J. & Kadonaga, J.T. (1991) Science, 254, 238–245.
5) Elgin, S.C. (1981) Cell, 27, 413–415.
6) Bernstein, B.E., Meissner, A., & Lander, E.S. (2007) Cell, 128, 669–681.
7) Rosenfeld, M.G., Lunyak, V.V., & Glass, C.K. (2006) Genes Dev., 20, 1405–1428.
8) Yokoyama, A., Fujiki, R., Ohtake, F., & Kato, S. (2011) Cold Spring Harb. Symp. Quant. Biol., 76, 165–173.
9) Deato, M.D. & Tjian, R. (2007) Genes Dev., 21, 2137–2149.
10) Goodrich, J.A. & Tjian, R. (2010) Nat. Rev. Genet., 11, 549–558.
11) Shi, Y., Lan, F., Matson, C., Mulligan, P., Whetstine, J.R., Cole, P.A., Casero, R.A., & Shi, Y. (2004) Cell, 119, 941–953.
12) Lee, M.G., Wynder, C., Cooch, N., & Shiekhattar, R. (2005) Nature, 437, 432–435.
13) Yokoyama, A., Igarashi, K., Sato, T., Takagi, K., Otsuka, I.M., Shishido, Y., Baba, T., Ito, R., Kanno, J., Ohkawa, Y., Morohashi, K., & Sugawara, A. (2014) J. Biol. Chem., 289, 18152–18162.
14) Kim, J.G. & Hudson, L.D. (1992) Mol. Cell. Biol., 12, 5632–5639.
15) Vierbuchen, T., Ostermeier, A., Pang, Z.P., Kokubu, Y., Südhof, T.C., & Wernig, M. (2010) Nature, 463, 1035–1041.
著者紹介Author Profile
 横山 敦(よこやま あつし)
横山 敦(よこやま あつし)東北大学大学院医学系研究科助教.農学博士.
略歴1979年群馬県に生まれる.2003年東京大学農学部卒業.09年同大学院農学生命科学研究科博士課程修了.07~09年日本学術振興会特別研究員(DC2).ERATO研究員,東京大学特任助教を経て現職.
研究テーマと抱負研究テーマは「新たな転写制御メカニズムの探索」.生化学的手法とプロテオミクスを組み合わせて転写関連因子の結合因子や翻訳後修飾の同定を行っています.膨大なMSスペクトルデータの中に未知の転写制御因子が埋もれていると信じています.
ウェブサイトhttp://www.med.tohoku.ac.jp/org/health/84/
趣味麺類.