二光子(多光子)励起顕微鏡を用いた蛍光生体イメージングは,①細胞レベルの高解像度で生体組織の断層像撮影ができる,②多色イメージングができる,③目的に応じた遺伝子組換えマウスを利用できる,といった利点がある.本稿では,目的タンパク質の活性を測定するFRET(蛍光共鳴エネルギー移動)バイオセンサーの原理と,FRETバイオセンサーを発現する遺伝子組換えマウスを用いた生体イメージング例を紹介し,二光子励起顕微鏡を用いた生体蛍光イメージングの技術的な課題などを紹介する.
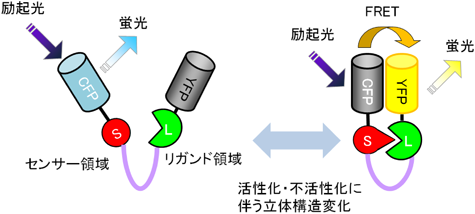
二つの蛍光分子が近接しているときに,励起された蛍光分子(ドナー分子)のエネルギーが他の蛍光分子(アクセプター分子)に遷移し,アクセプター分子が蛍光を発する現象を「蛍光共鳴エネルギー移動」(fluorescence resonance energy transfer: FRET)と呼ぶ.頭文字のFはfluorescenceの代わりに発見者フェルスターの名前をとってFörster resonance energy transferと呼ぶ場合もある.このFRETの原理を利用して,タンパク質分子間の距離,構造変化,酵素活性を検出するためのさまざまなバイオセンサー(FRETバイオセンサー)が開発されている1,2).一般的なFRETバイオセンサーはシアン色蛍光タンパク質(CFP)をドナー分子に,黄色蛍光タンパク質(YFP)をアクセプター分子にしてさらに,センサー領域,リガンド領域を有する構造を持つ(図1).センサー領域は外部の環境変化を捉える領域で,リン酸化ドメイン,GTP結合ドメインなどで構成される.リガンド領域はセンサー領域の構造変化に対して結合または解離するようなタンパク質ドメインが用いられる.センサー領域とリガンド領域を一分子に連結させた「一分子型」FRETバイオセンサーでは,センサー領域,リガンド領域をFRETに適した位置にデザインする必要がある3).
3. FRETバイオセンサーを発現する遺伝子組換えマウスの作出
FRETバイオセンサーを発現する遺伝子組換えマウス(以下,FRETマウス)を作出するためには,生体内で長期間,安定的にFRETバイオセンサーを発現させる必要がある.しかし,ともにクラゲ由来であるCFP遺伝子とYFP遺伝子には高い相同性があるため,レトロウイルス,レンチウイルスを用いた遺伝子導入法では逆転写反応中に相同組換え反応が起こってしまう.また,直鎖上のDNAをゲノムに挿入させる方法は導入効率が悪く,導入遺伝子がサイレンシングを受けることが多い.東京大学の山口,三浦らは導入遺伝子を安定化させるインスレーターを用いて,カスパーゼ活性を測るFRETマウスの作出に成功し,胚での神経管閉鎖時におけるアポトーシス過程をイメージングした4).また,WelchらはROSA26遺伝子座に低分子量Gタンパク質Rac1のFRETバイオセンサー遺伝子を導入し,FRETマウスの作出を行った5).この他にもFRETマウスの作出に関する報告はあるが,十分な蛍光強度を持つマウス個体をイメージングしている例は数少ない6).筆者らはTol2トランスポゾンを利用した遺伝子導入方法を用いて,リン酸化酵素ERK(extracellular signal-regulated kinase)およびPKA(protein kinase A)の活性をイメージングできるFRETマウスを簡便に,高効率で作出することに成功した7).いずれの方法でもFRETマウスを用いた生体蛍光イメージングには,バイオセンサーの低毒性,標的組織での十分な発現(蛍光輝度)などが求められる(「5. 二光子励起顕微鏡による生体イメージングの実際」も参照のこと).
筆者らはリン酸化酵素ERKおよびPKAの活性をモニターするFRETマウスを用いて,耳の表皮から真皮の軟骨組織近くまでの厚さ約200 µmを二光子顕微鏡で観察した.ERKは細胞増殖をつかさどるシグナル伝達システム,MAPキナーゼ経路(Ras-Raf-MEK-ERK)の下流にあるリン酸化酵素であり,このMAPキナーゼ経路の異常な活性化は細胞のがん化を引き起こすことが知られている.またホルモンなどの刺激は細胞膜の受容体の活性化を経て,細胞内のセカンドメッセンジャー,サイクリックAMPを介してPKAを活性化する.活性化したPKAはさまざまな基質タンパク質をリン酸化することでシグナルを下流に伝達する.マウスの耳の皮膚は心臓や肺から離れており,拍動の影響をほとんど受けないため,最も生体イメージングしやすい部位の一つである.皮膚の基底細胞に注目すると,PKAの活性はどの細胞でもほぼ同じであるのに対して,ERKの活性は細胞ごとにばらついていた.レーザーを集中的に照射して基底細胞層の一部を融除させ,その周囲で起こるPKAとERKの活性変化を調べたところ,レーザーで融除した領域近傍の基底細胞ではPKAとERKはともに活性化し,その活性化状態は少なくとも1時間続いた.皮膚などの生体組織が損傷を受けると直ちに炎症,免疫応答,組織修復を促す分子種「アラーミン」が放出されることが知られている.レーザー照射によって融除領域から熱やアラーミンが放出され,周囲の細胞に創傷治癒を促すシグナルが伝わった結果,PKA,ERKの活性化が起こったと推測できる.
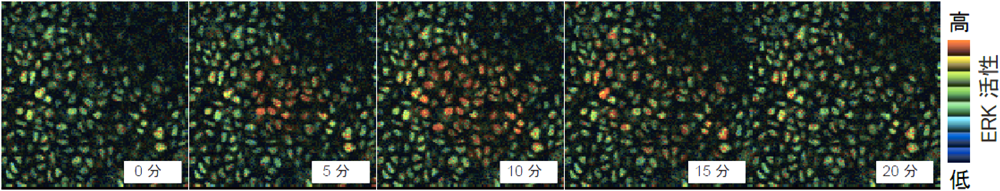
また当研究室の平塚らは,傷の修復過程にある皮膚を長時間にわたって観察することで,ERKの活性化が同心円状に広がっていく現象を発見し,SPREAD(spatial propagation of radial ERK activity distribution)と名づけた(図2).SPREADの発生メカニズムを調べたところ,表皮中に存在する細胞増殖因子がERKの活性化により切断されて隣の細胞を活性化し,その細胞の中で,またERKが活性化されるという連鎖反応が起こっていた8).
当研究室の水野らは,FRETマウスの小腸組織における好中球の血管外遊走メカニズムを詳細に解析した9).マウス小腸の絨毛基部にはクリプト(陰窩)と呼ばれる「くぼみ」がある.マウス小腸を体外へ露出させ,血流を保ちながら顕微鏡ステージ上で動かないようにして二光子励起顕微鏡観察を行うと,クリプトから絨毛までを連続撮影することができる(図3).また,クリプト周囲には血管が豊富に存在するため,腸管内にリポ多糖(LPS)などの炎症誘発物質を入れることで,好中球遊走モデルを作製することができる.好中球の血管外遊走メカニズムに関しては古くから調べられており,一連の細胞接着反応の中で,ERKがインテグリンなどの接着分子の活性を制御するInside-out signalと,逆に接着分子がERKの活性を制御するOutside-in signalが働くことが報告されている.しかし,実際に一連の血管外遊走プロセスにおいて好中球のERK活性がどのように変化するのかはこれまで確かめられたことはなかった.FRETマウスの生体イメージングによって,①ERK活性化は好中球が血管内皮細胞に接着した段階から上昇すること,②ERK活性は間質に浸潤した後も高いままであること,③生理活性脂質LTB4が腸管に浸潤した好中球のERK活性を上昇させること,④間質ではPKAの活性が好中球の運動速度と逆相関を示すこと,⑤PKAはERK活性を抑制し,細胞運動を阻害すること,⑥非ステロイド性抗炎症薬(NSAIDs)はERKを活性化し,好中球の集簇を増強することなどがわかった.炎症部から遠いところではLTB4-BLT1受容体シグナルの下流で活性化する三量体Gタンパク質Giの下流でERKが優位となり,好中球の遊走が促進される.一方,炎症中心部ではPGE2-EP4受容体シグナルの下流で三量体Gタンパク質Gsが優位となり,PKAが活性化することで好中球の遊走を抑制するシグナルが働くらしい.
5. 二光子励起顕微鏡による生体イメージングの現状と課題
生体組織は厚みのある散乱媒体であるため,細胞レベルの光学断層撮影に用いられる共焦点顕微鏡では生体組織の蛍光イメージングは難しい.そこで,生体組織を蛍光イメージングする場合には,二光子励起顕微鏡がよく用いられる.二光子励起とは,一つの蛍光分子が二つの光子を同時に吸収して励起状態になる非線形光学現象である.二光子励起には,通常の(一光子励起の場合の)2倍の吸収波長の光子を焦点に高密度に集める必要があるため,近赤外領域のパルスレーザー光が利用される.この近赤外領域の光は組織透過性が高く,生体組織のイメージングに向いている.二光子励起現象は焦点でのみ起こるため,蛍光分子の褪色や光毒性の影響が少ないことも二光子励起顕微鏡の大きなメリットである.ただし,ここで注意しておくべき点は二光子励起顕微鏡で組織の深部観察が可能といっても,せいぜい組織表面から数百µmから1 mmほどの深さまでであり,さらに観察可能な深さは組織・部位ごとに異なる点である.これは次にあげる自家蛍光の問題とも深く関わっている.
蛍光イメージングでの難題の一つは「生体組織由来の自家蛍光」である.特に,使用する蛍光プローブやバイオセンサーの蛍光強度が弱い場合に,組織由来の自家蛍光は無視できない.生体内にはNAD(P)H,FAD,脂質(リポフスチン)などの自家蛍光物質が存在するが,これらを生体中から除くことはできない.そのため,十分に明るい蛍光物質を利用する,自家蛍光物質を励起しないレーザー波長を用いる,などの工夫が必要になる.しかし逆に,組織由来の自家蛍光をうまく利用して無染色で生体イメージングすることも可能である10,11).
二光子励起顕微鏡を用いてマウスの生体イメージングを行う際のさらに難しい課題は「組織の固定(不動化)」である.特に心臓の拍動や呼吸による動きを観察部位から除くことは難しい場合が多い.周期的に観察視野が動いている場合は,解析時にソフトウエア上である程度補正することが可能である.組織の固定には,物理的に圧迫して固定する方法が簡便であるが,抑えすぎると血流が止まり,組織が壊死してしまう.また固定のための圧迫や乾燥の影響で,時間とともに組織の厚みが変わってしまうことも長時間のイメージングの際には問題になることが多い.
近年,日本でもマウス生体イメージングに関する書籍が出版され12),さまざまな情報交換の場が形成されているので,こういったものを活用して問題解決していくこともおすすめである13,14).
CRISPR/Casシステムなどのゲノム編集技術の進歩により,マウス個体を用いた実験系が生命科学の発展にますます重要になることが予想できる.FRETマウスの種類も増え,新しいイメージング手法が開発されることによって,より多くの生命現象がin vivoでみえてくるであろう.我々は,これまで実験に貢献してもらったマウスに感謝の意を表し,今後もFRETマウスが生体内環境をイメージングするための優れた研究材料になることを期待している.
謝辞Acknowledgments
本稿で紹介した研究の内容は,京都大学大学院医学研究科病態生物医学(松田道行教授)講座で実施されたものです.長年にわたるご指導と研究室メンバーのご協力に深く感謝いたします.
引用文献References
1) Miyawaki, A. & Niino, Y. (2015) Mol. Cell, 58, 632–643.
2) Kiyokawa, E., Aoki, K., Nakamura, T., & Matsuda, M. (2011) Annu. Rev. Pharmacol. Toxicol., 51, 337–358.
3) Komatsu, N., Aoki, K., Yamada, M., Yukinaga, H., Fujita, Y., Kamioka, Y., & Matsuda, M. (2011) Mol. Biol. Cell, 22, 4647–4656.
4) Yamaguchi, Y., Shinotsuka, N., Nonomura, K., Takemoto, K., Kuida, K., Yosida, H., & Miura, M. (2011) J. Cell Biol., 195, 1047–1060.
5) Johnsson, A.K., Dai, Y., Nobis, M., Baker, M.J., McGhee, E.J., Walker, S., Schwarz, J.P., Kadir, S., Morton, J.P., Myant, K.B., Huels, D.J., Segonds-Pichon, A., Sansom, O.J., Anderson, K.I., Timpson, P., & Welch, H.C. (2014) Cell Reports, 6, 1153–1164.
6) Jacobs, S., Calebiro, D., Nikolaev, V.O., Lohse, M.J., & Schulz, S. (2010) Endocrinology, 151, 4560–4565.
7) Kamioka, Y., Sumiyama, K., Mizuno, R., Sakai, Y., Hirata, E., Kiyokawa, E., & Matsuda, M. (2012) Cell Struct. Funct., 37, 65–73.
8) Hiratsuka, T., Fujita, Y., Honda, N., Aoki, K., Kamioka, Y., & Matsuda, M. (2015) eLife. 10.7554/eLife.05178
9) Mizuno, R., Kamioka, Y., Kabashima, K., Imajo, M., Sumiyama, K., Nakasho, E., Ito, T., Hamazaki, Y., Okuchi, Y., Sakai, Y., Kiyokawa, E., & Matsuda, M. (2014) J. Exp. Med., 211, 1123–1136.
10) Orzekowsky-Schroeder, R., Klinger, A., Martensen, B., Blessenohl, M., Gebert, A., Vogel, A., & Huttmann, G. (2011) J. Biomed. Opt., 16, 116025.
11) Vielreicher, M., Schürmann, S., Detsch, R., Schmidt, M.A., Buttgereit, A., Boccaccini, A., & Friedrich, O. (2013) J. R. Soc. Interface, 10, 20130263.
12) 石井優編(2012)in vivoイメージング実験プロトコール(実験医学別冊最強のステップUPシリーズ),羊土社.
13) 新学術領域研究「蛍光生体イメージ」 http://www.imaging.lif.kyoto-u.ac.jp/
14) Bioimage.jp http://www.bioimage.jp/
著者紹介Author Profile
 上岡 裕治(かみおか ゆうじ)
上岡 裕治(かみおか ゆうじ)京都大学大学院医学研究科特定助教.博士(医学).
略歴1976年大阪府に生る.99年大阪大学工学部応用生物工学科卒業.2001年大阪大学大学院工学研究科修士課程修了.企業への就職を経て,07年大阪大学大学院医学系研究科博士課程修了.08年日本学術振興会特別研究員PD. 同年京都大学大学院生命科学研究科特定研究員,京都大学大学院医学研究科特定助教.
研究テーマと抱負生体イメージングによる癌微小環境の解明に取り組んでいます.