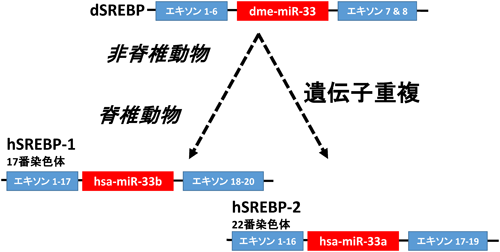
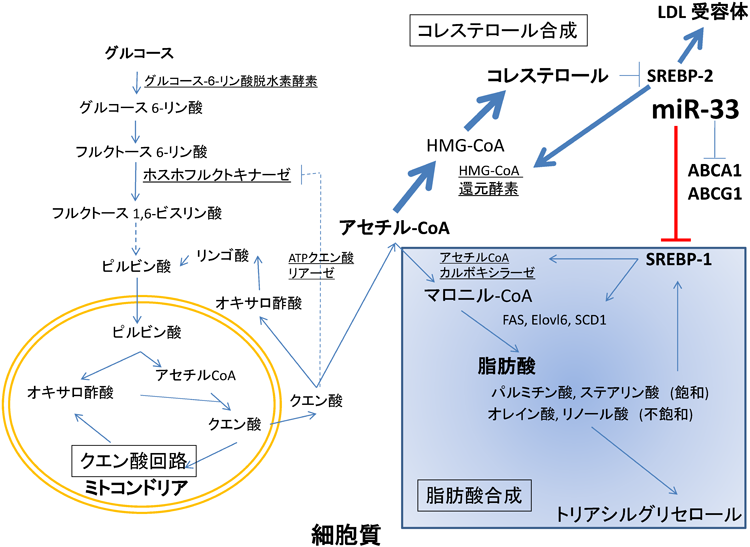
3) Seegmiller, A.C., Dobrosotskaya, I., Goldstein, J.L., Ho, Y.K., Brown, M.S., & Rawson, R.B. (2002) Dev. Cell, 2, 229–238.
5) Grimson, A., Srivastava, M., Fahey, B., Woodcroft, B.J., Chiang, H.R., King, N., Degnan, B.M., Rokhsar, D.S., & Bartel, D.P. (2008) Nature, 455, 1193–1197.
6) Rayner, K.J., Suarez, Y., Davalos, A., Parathath, S., Fitzgerald, M.L., Tamehiro, N., Fisher, E.A., Moore, K.J., & Fernandez-Hernando, C. (2010) Science, 328, 1570–1573.
7) Najafi-Shoushtari, S.H., Kristo, F., Li, Y., Shioda, T., Cohen, D.E., Gerszten, R.E., & Naar, A.M. (2010) Science, 328, 1566–1569.
8) Horie, T., Ono, K., Horiguchi, M., Nishi, H., Nakamura, T., Nagao, K., Kinoshita, M., Kuwabara, Y., Marusawa, H., Iwanaga, Y., Hasegawa, K., Yokode, M., Kimura, T., & Kita, T. (2010) Proc. Natl. Acad. Sci. USA, 107, 17321–17326.
9) Horie, T., Baba, O., Kuwabara, Y., Chujo, Y., Watanabe, S., Kinoshita, M., Horiguchi, M., Nakamura, T., Chonabayashi, K., Hishizawa, M., Hasegawa, K., Kume, N., Yokode, M., Kita, T., Kimura, T., & Ono, K. (2012) J. Am. Heart Assoc, 1, e003376.
10) Horie, T., Nishino, T., Baba, O., Kuwabara, Y., Nakao, T., Nishiga, M., Usami, S., Izuhara, M., Sowa, N., Yahagi, N., Shimano, H., Matsumura, S., Inoue, K., Marusawa, H., Nakamura, T., Hasegawa, K., Kume, N., Yokode, M., Kita, T., Kimura, T., & Ono, K. (2013) Nat. Commun., 4, 2883.
11) Horie, T., Nishino, T., Baba, O., Kuwabara, Y., Nakao, T., Nishiga, M., Usami, S., Izuhara, M., Nakazeki, F., Ide, Y., Koyama, S., Sowa, N., Yahagi, N., Shimano, H., Nakamura, T., Hasegawa, K., Kume, N., Yokode, M., Kita, T., Kimura, T., & Ono, K. (2014) Sci. Rep, 4, 5312.