1. はじめに:細菌のリボソーム解放システム,トランストランスレーション
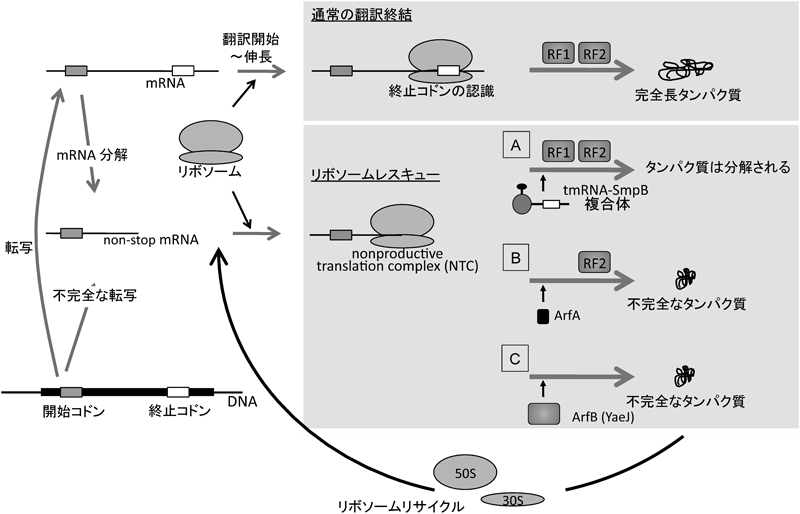
4種類のヌクレオチドで記述される遺伝情報は,連続する三つのヌクレオチドが一組のコドンとして一つのアミノ酸を指定し,タンパク質の一次構造を決定する.43=64種類のコドンのうち61種類はアミノ酸を指定するが,3種類は指定するアミノ酸を持たない,いわゆる終止コドンである.終止コドンは単に「対応するアミノアシルtRNAが存在しない」コドンではなく,「翻訳終結因子(release factor: RF)によって認識される」コドンであり,RFによるペプチジルtRNA加水分解に始まる一連の翻訳終結反応のシグナルとして積極的な役割を担う.また,終止コドンは開始コドンとともにオープンリーディングフレーム(open reading frame: ORF)を規定する.終止コドンがなければ翻訳産物のC末端が定まらないという事態に陥る.すなわち,下世話な言葉でいえば「終止コドンは大事」なのである.では,終止コドンがない場合,翻訳はどうなるのであろうか.終止コドンを持たないnon-stop mRNAであっても,SD(Shine-Dalgarno)配列と開始コドンがそろっていれば翻訳は開始される.しかし,その場合リボソームはmRNAの末端に到達しても正常に翻訳を終結できずに立ち往生し,mRNA,ペプチジルtRNAなどからなる複合体,nonproductive translation complex(NTC)が形成される(図1).
細菌はトランストランスレーション(trans-translation)と呼ばれる特殊な翻訳機構によって立ち往生したリボソームを解放してNTCを解消することが知られる(図1A)1–4).trans-translationに関してはさまざまな総説(たとえば本誌79巻,姫野ら4))が出版されているため,ここで詳細な解説は控えるが,ごく簡単に表現すれば,ssrA遺伝子にコードされるtmRNA(SsrA RNA)分子がnon-stop mRNAに継ぎ足される形で翻訳が続行し,tmRNA上の終止コドンで翻訳が終結することでNTCが解消されるシステムである2–5).継ぎ足された部分のRNAも当然ながら翻訳され,そこにコードされるペプチド配列がC末端に融合されたポリペプチド鎖ができ上がる.このC末端に融合される配列はSsrAタグと呼ばれ,SsrAタグがC末端に付加したタンパク質はプロテアーゼにより積極的に分解される1–4).
NTCの蓄積は,リボソームの実効濃度の減少による細胞全体の翻訳活性の低下を招き,有害であると考えられる.trans-translationはNTCの解消を通し,翻訳活性を維持する効果をもたらす.さらに,non-stop mRNAにコードされる不完全なポリペプチドは機能しないだけでなく,むしろ有害である可能性もあるため,産物が分解されるのは理にかなっている.また,non-stop mRNAの3′末端に形成されるNTCはヌクレアーゼのアクセスに対する障害となることからNTCの解消はnon-stop mRNAを減少させることにもなる.このようにtrans-translationは翻訳活性の維持,タンパク質品質管理,mRNA品質管理と,複数の重要な効果をもたらす機構として認識されている2–4).
最近,大腸菌においてtrans-translationのバックアップ機構とでもいうべきリボソーム解放機構が発見され,リボソーム解放の重要性があらためて示された.以下でその新たなリボソーム解放機構について,その発見の経緯,巧妙な発現制御,多彩な分子機構を概説したい.
昨今,盛んにゲノム解析が行われ,さまざまな真正細菌のゲノム情報が明らかになってきている.それによると,ssrAは,ごく少数の例外を除きすべての真正細菌で見いだされている.加えて,NTCの解消と不完全なポリペプチド鎖の除去という機能を担うことから,trans-translationは細菌の生命維持において重要であると考えられた.実際,淋菌,マイコプラズマ,ピロリ菌等ではtmRNAが生育に必須であるということを示す(または示唆する)報告がなされている2,3).しかし一方で,大腸菌,枯草菌等多くの細菌でtmRNA欠損株が得られ,必ずしもこれらの菌株ではtrans-translationは必要ではないらしいことも示されており,trans-translationの生理学的な存在意義に関しては疑問が残されていた.
そこで我々は,大腸菌のtmRNAが必須でないのはtrans-translationにかわる第二のNTC解消機構が存在するからであり,その因子を欠く大腸菌株は生育にtmRNAを必要とする,との作業仮説のもと,その第二の因子の探索を試みた.その結果,機能不明のORF,yhdLに塩基置換が生じた株が得られた6).yhdLは大腸菌ゲノム上で72アミノ酸からなるタンパク質をコードしうる(わざわざこのような回りくどい表現をするのには後述のような理由がある)ORFであり,得られた変異株ではその18番目のトレオニンがアラニンに置換していた.
yhdL欠損株は通常は何の表現型も示さないが,ssrAの欠損や,SsrAとともにtrans-translationに必須の遺伝子smpBの欠損と併せると合成致死の表現型を示す.すなわちyhdLはtrans-translationができない大腸菌の生育には必須である.興味深いことに,yhdLとssrAの合成致死の表現型は亜致死濃度のピューロマイシンにより,部分的に抑圧される.ピューロマイシンはアミノアシルtRNAのミミックとしてペプチド基を受け取り,その時点で翻訳を終結させる抗生物質であり,その性質上NTCに作用すればそれを解消しうる.これはすなわち,yhdL ssrA二重欠損が生育できないのはNTCの蓄積が原因である,と解釈でき,最初の仮説どおりyhdLがtrans-translationにかわる第二のNTC解消因子であることを強く示唆する結果である.そこで,明確な証拠を得るために,ORF内に転写ターミネーターを挿入したモデル遺伝子を細胞内で発現させた.転写ターミネーターの作用で終止コドンに到達する前に強制的に転写が終結されるため,この遺伝子が発現するとnon-stop mRNAが生産され,その末端でNTCが形成される.ssrA,yhdLそれぞれの欠損や過剰発現を組み合わせたさまざまな条件でモデル遺伝子の産物を解析した結果,実際にyhdLがNTC解消に関与していることが示された.
さらに,精製したYhdLタンパク質がin vitroでNTC解消活性を示すことも明らかとなった.大腸菌yhdL欠損株の抽出液にSsrA RNAのタグコード領域に対合するオリゴヌクレオチドを加え,trans-translationを阻害することで,ssrA yhdL二重欠損の状態を再現できる.その状態で,人工合成したnon-stop mRNAを翻訳させると,NTCが蓄積する.NTCの蓄積はペプチジルtRNAの増加を指標に解析できる.そこに精製したYhdLを加えると,確かにペプチジルtRNAは減少し,YhdLタンパク質によるNTC解消がin vitroでも示された.以上の解析から,YhdLタンパク質がNTCを解消することが明らかとなった(図1B).これを受け,yhdLをalternative ribosome rescue factor Aという意味でarfAと改名することを提案し6),広く認められている.これ以降,現在の名称であるarfAを用いる.
arfA ssrA二重欠損株の示す合成致死のマルチコピーサプレッサーとしてyaeJが単離された7).yaeJの翻訳産物であるYaeJタンパク質は,ペプチジルtRNA加水分解に必須とされるいわゆるGly-Gly-Gln(GGQ)モチーフ8)を持つ翻訳終結因子のホモログである.YaeJのヒトミトコンドリアにおけるホモログICT-1タンパク質はNTC解放活性を持つことが報告されていたが9),YaeJが大腸菌で同様の活性を持つことは知られていなかった.筆者らのグループが遺伝学的解析からYaeJのNTC解放活性を明らかにした(図1C)のと同時期に,群馬大学の行木研究室では構造生物学的解析から同様の結論に達し10),それぞれ独立に報告している.我々は第三のNTC解消因子であることからyaeJをarfB(alternative ribosome rescue factor B)と改称することを提案している7).なお,arfA ssrA二重欠損株が合成致死を示すということは,染色体上のarfBは単独では細胞内で生じるNTCを生存可能なレベルまで減ずることはできないことを示す.
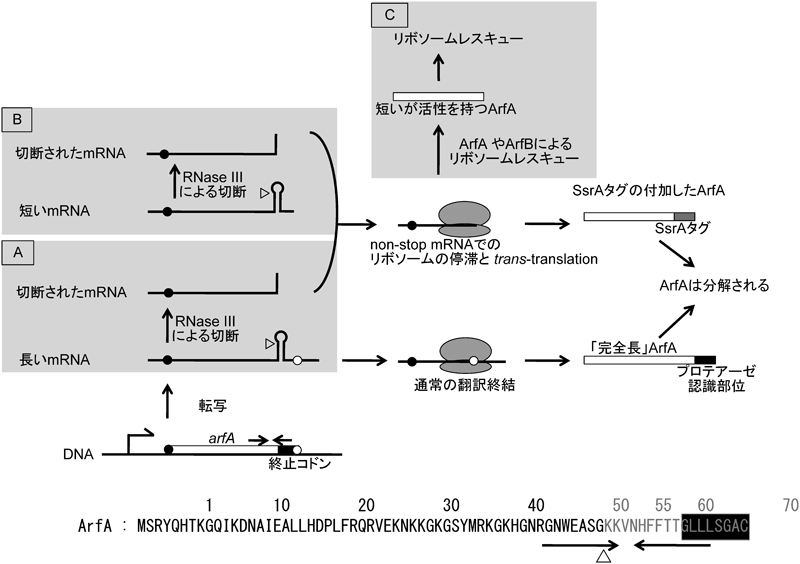
4. ArfAの発現制御:trans-translationによる発現抑制
ArfAの発現は,その機能と密接に関わる特徴的な制御を受けている(図2).完全長のArfAの細胞内での発現は低レベルで,arfA mRNAもほとんど検出されないが,ArfAのC末端を欠失させるとArfAの生産量が増加し,さらにmRNAも安定化する.その原因がarfA ORF内部に存在するステムループ構造をとりうる逆向き反復配列であることがわかり,arfAの特徴的な発現制御機構が明らかとなった11,12).arfA mRNA上のこの逆向き反復配列が形成するステムループ構造はRNase IIIによって認識,切断される.その結果,arfA mRNAはnon-stop mRNAとなる.前述のようにnon-stop mRNAの3′末端ではNTCが形成され,通常はtrans-translationによって解消される.同時に,作られかけていたポリペプチドはSsrAタグが付加されることで分解へと導かれる.arfAの場合も例外ではなく,「後半部分を失ったArfA」は分解される.このため完全長のArfAの細胞内での発現は低レベルであった.RNase IIIの欠損や,ステムループ構造をとらせない(アミノ酸配列には影響しない)ようにarfAに導入した塩基置換変異はいずれもArfA産生量の増加につながった.この結果からArfAの独特の発現制御機構を描くことができた.すなわち,arfAは通常はnon-stop mRNAとして発現し,trans-translationの標的となって分解され続けるため,細胞内には(あるとしても)ほとんど存在しない.しかし,何らかの要因で細胞内のnon-stop mRNAが増加し,trans-translationシステムの処理能力を超えたとき,分解を免れた(C末端を欠く)ArfAがNTC解消因子としてその機能を発揮する,という機構である.これはtrans-translationのバックアップ機構としてのArfAの発現制御機構としては,大変理にかなっている.trans-translationはNTCを解消すると同時に,潜在的に有害である不完全なポリペプチドを分解へと導く.それに対してArfAはNTCを解消するものの,ポリペプチドは分解され残る.細胞内システムの健全性を維持するためには,trans-translationの方が細胞にとって「安全・確実」であると思われる.ただし,NTCの蓄積がtrans-translationの処理能力を超えた場合,(ポリペプチドは分解され残るものの)ArfAの助けも必要となる.まさにそのような状況でArfAは分解を免れて発現し,NTC解消因子としての機能を発揮するのであろう.RNase IIIによりarfA mRNAは55番目のコドン付近で切断される.実際,ssrA変異株では56番目以降のアミノ酸を欠失するArfAが検出できるのだが,人為的に56番目以降のアミノ酸を欠失させたArfA,すなわち上記の機構で発現するArfAはNTC解放活性を保持しており,このシナリオと矛盾しない.翻訳を停止させるストレス,たとえば抗生物質の添加によりArfAの発現の増加が観察されることもこれを支持する.
trans-translationが関わるArfAの発現制御は大変厳密である.転写産物の解析からarfAの転写は逆向き反復配列のすぐ下流で終結することが示されたが,この転写終結点もarfA ORFの内部に存在し,たとえRNase IIIによる分解を免れても大部分のarfA mRNAはnon-stop mRNAとなる.さらに,仮に転写がarfAのゲノム上のORFをカバーするまでに進み得たとしても,その(ゲノム上にコードされたとおりの)翻訳産物はC末端付近の疎水性アミノ酸からなるクラスターがプロテアーゼの標的となることから大変不安定で,速やかに分解される.つまり,ArfAは転写終結,mRNAプロセシング,タンパク質分解の三重の発現抑制機構により,事実上trans-translationシステムの処理能力を超えたnon-stop mRNAが存在するときにのみ発現するように制御されているのである.
さらに,このArfAの発現制御機構はArfBの存在意義についても説明できる.上述のようにarfA ssrA二重欠損が致死であるということ6)は,大腸菌ゲノム上のarfBは,少なくともこの状態においては大腸菌の生育を維持できないことになる.つまり,ArfBは,ArfA,SsrA双方が存在しないという(通常生育している大腸菌はめったに経験することのない)特殊な状況で生育を支える因子であるとは考えにくい.むしろ,ArfAが必要とされる状況,すなわち,non-stop mRNAが危険なまでに蓄積した状況で,より早くArfAを発現させ,NTCを解消させるという役割を担っているのではないだろうか.あまりにも厳密にその発現が制御されているため,ArfAは,必要とされる状況になってもすぐには発現できない恐れがあるが,ArfBはそのレスポンスを少しでも早めることができる.ArfA,ArfBはtrans-translationと協働して,細胞の健全性を維持するNTC解消システムを構成しているのであろう.
このArfAの発現制御系は,もう一つ,大きな意味を持つ.すなわち,arfAは,non-stop mRNAとして発現するようプログラムされ,しかもそれがその機能と密接に関わっているこれまで知られている唯一の遺伝子なのである.
自らGGQモチーフを備えるArfBと異なり,実質55アミノ酸残基程度のArfAが単独でNTCを解消できるとは考えにくい.むしろRFなどの他の因子と協働していると考えるのが自然であろう.実際,ArfAは単独ではNTCを解消できず,in vitro系での探索の結果,ArfAによるNTC解消にはRF2が必要であることが明らかとなった13,14).おそらく,何らかの機構でNTCを認識したArfAがRF2をリクルートし,そのペプチジルtRNA加水分解活性によりNTCを解消させるのであろう.面白いことに終止コドンの認識を改変したRF2でも結果は同じであった.一方でRF1はこの機構には関与しなかった.RF1とRF2は認識配列の一部を共有し,同じようにペプチジルtRNAを加水分解するが,決して同一の因子ではない.ArfAとRF2の関係は,RF1,RF2の起源に関しても何らかのヒントを与えてくれるかもしれない.
ArfAによるNTCの解消にRF2が要求されることから,もう一つ導き出されるルールがある.それはリボソーム内のペプチジルtRNAを加水分解するには,RF,またはそのホモログであるArfBのGGQモチーフが必須である,というルールである.
ArfAがNTCを認識し,おそらくはNTCに結合し,RF2をリクルートすることでNTCを解消するという構図は描けた.しかし,どのように認識するのか,どのように結合するのか,どのようにリクルートするのか,その具体的な分子機構は依然として不明である.最近,弘前大の姫野研究室がArfAとRF2のNTC内における挙動の手がかりを見いだした15).ヒドロキシラジカルプロービング法でリボソーム内のArfAの位置を解析したところ,AサイトのデコーディングセンターとmRNAエントリーチャンネルのごく近傍に位置し,そこにRF2が加わると結合様式が変化することが示された.これは,ArfAがNTCを認識し,リボソームAサイトに結合し,次いでRF2をリクルートするというモデルを支持する結果である.ArfAとNTC中のリボソームとの結合をより詳細に解析することで,ArfAとRF2によるNTC解消の分子機構の詳細を明らかにするのが現在の課題である.
複製,転写,翻訳,いずれの反応も,その開始段階が重要であることは,開始しなければ何も起こらないことからも明らかである.しかし,これら情報高分子の合成反応は,その終結段階も重要であり,軽視してよいものではないということが,この研究を行っていてあらためて認識された.
細胞内でのmRNAのターンオーバーは活発であり,これが遺伝子発現の迅速なオン・オフの調節を保証している.しかし,mRNAはその分解の過程で必ず終止コドンを持たない状態を経験する.すなわち,non-stop mRNAは細胞内で常に一定量存在しており,必然的にNTCも常に形成されている.細菌は大きなエネルギーを消費してでもこのNTCの蓄積を避け,翻訳系の健全さを保証している.意外と知られていないこの「終わりの保証」にもさまざまな面白さが潜んでいる.
謝辞Acknowledgments
岡山大学で同じ研究グループとしてさまざまな面で支えて下さる沓掛和弘教授,冨永晃准教授,これまで在籍した学生,研究員の皆さん,さらにこれまでお世話になった多くの方々にあらためて御礼申し上げたい.
引用文献References
1) Keiler, K.C., Weller, P.R., & Sauer, R.T. (1996) Science, 271, 990–993.
2) Abo, T. & Chadani, Y. (2014) Front. Microbiol, 5, 156.
3) Himeno, H., Nameki, N., Kurita, D., Muto, A., & Abo, T. (2015) Biochimie, 114, 102–112.
4) 姫野俵太,栗田大輔,高田一馬,今野貴之,塙(末次)京子,竹本千重,川添将仁,横山茂之,行木信一,河合剛太,武藤昱(2007)生化学,79, 213–221.
5) Karzai, A., Susskind, M., & Sauer, R. (1999) EMBO J., 18, 3793–3799.
6) Chadani, Y., Ono, K., Ozawa, S., Takahashi, Y., Takai, K., Nanamiya, H., Tozawa, Y., Kutsukake, K., & Abo, T. (2010) Mol. Microbiol., 78, 796–808.
7) Chadani, Y., Ono, K., Kutsukake, K., & Abo, T. (2011) Mol. Microbiol., 80, 772–785.
8) Frolova, L.Y., Tsivkovskii, R.Y., Sivolobova, G.F., Oparina, N.Y., Serpinsky, O.I., Blinov, V.M., Tatkov, S.I., & Kisselev, L.L. (1999) RNA, 5, 1014–1020.
9) Richter, R., Rorbach, J., Pajak, A., Smith, P.M., Wessels, H.J., Huynen, M.A., Smeitink, J.A., Lightowlers, R.N., & Chrzanowska-Lightowlers, Z.M. (2010) EMBO J., 29, 1116–1125.
10) Handa, Y., Inaho, N., & Nameki, N. (2011) Nucleic Acids Res., 39, 1739–1748.
11) Chadani, Y., Matsumoto, E., Aso, H., Wada, T., Kutsukake, K., Sutou, S., & Abo, T. (2011) Genes Genet. Syst., 86, 151–163.
12) Garza-Sánchez, F., Schaub, R.E., Janssen, B.D., & Hayes, C.S. (2011) Mol. Microbiol., 80, 1204–1219.
13) Chadani, Y., Ito, K., Kutsukake, K., & Abo, T. (2012) Mol. Microbiol., 86, 37–50.
14) Shimizu, Y. (2012) J. Mol. Biol., 423, 624–631.
15) Kurita, D., Chadani, Y., Muto, A., Abo, T., & Himeno, H. (2014) Nucleic Acids Res., 42, 13339–13352.
著者紹介Author Profile
 阿保 達彦(あぼ たつひこ)
阿保 達彦(あぼ たつひこ)岡山大学大学院自然科学研究科(理学部生物学科)准教授.博士(農学).
略歴1965年東京都に生る(親が転勤族だったため実質的には埼玉県出身).89年東京大学農学部農芸化学科卒業.94年同大学院農学系研究科農芸化学専攻修了.98年名古屋大学大学院理学研究科助手.2002年より現職.
研究テーマと抱負健全な翻訳系の維持のしくみを明らかにする.
ウェブサイトhttp://www.biol.okayama-u.ac.jp
趣味読書,旅(計画,実行,妄想).