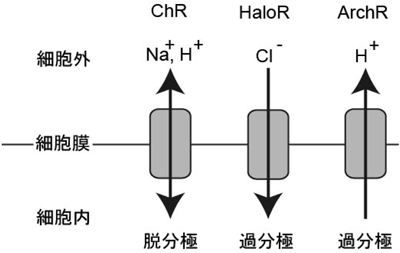
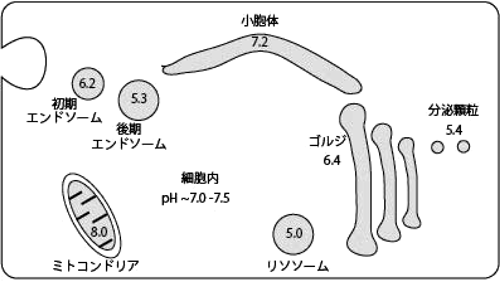
オプトジェネティクスに期待される非興奮性細胞の新たな操作技術Optogenetics on non-excitable cells
慶應義塾大学医学部精神・神経科学教室Department of Neuropsychiatry, Keio University School of Medicine ◇ 〒160–8582 東京都新宿区信濃町35 ◇ 35 Shinanomachi, Shinjuku, Tokyo 160–8582, Japan
発行日:2016年4月25日Published: April 25, 2016