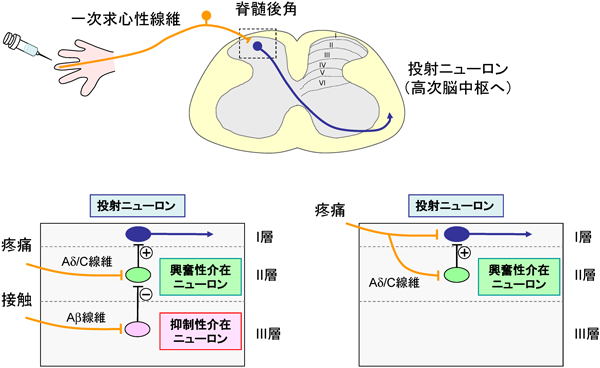
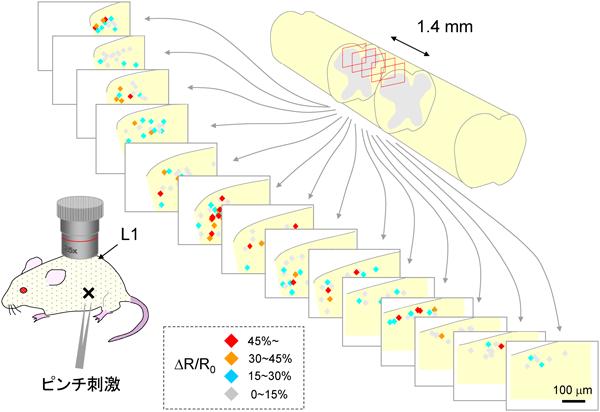
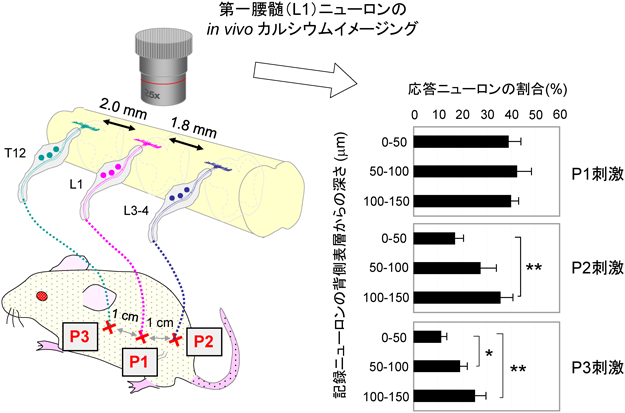
脊髄後角神経回路による体性感覚の情報処理Neuronal circuitry for sensory processing in the spinal dorsal horn
関西医科大学医化学講座Department of Medical Chemistry, Kansai Medical University ◇ 〒573–1010 大阪府枚方市新町2–5–1 ◇ 2–5–1, Shinmachi Hirakata, Osaka 573–1010, Japan
発行日:2016年4月25日Published: April 25, 2016