1)概要
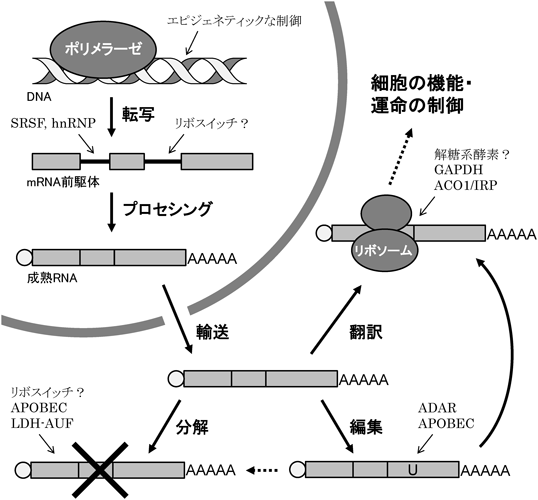
転写された直後のRNAは一次転写産物と呼ばれる状態であり,そのうちタンパク質をコードするものがいわゆるmRNA前駆体(pre-mRNA)である.pre-mRNAはそのままではタンパク質の翻訳に使用されず,塩基修飾やスプライシングなどのプロセシングを経て,初めて翻訳が可能な成熟mRNAとなる(図1).すなわちRNAのプロセシングは,タンパク質発現の調節における重要な制御点である.RNAプロセシングおよび翻訳の過程における栄養素や中間代謝産物による影響は以前から知られていたが,近年,そのメカニズムがより詳らかにされつつある.また,代謝産物そのものがRNAに結合してその動態を制御する「リボスイッチ(riboswitch)」様の制御機構が,ヒトゲノムにも存在しうることが示唆されている.
なお,DNAと同様に,RNAはメチル化修飾されることによってもプロセシングや翻訳の調節を受ける.したがってメチル基供与体となるS-アデノシルメチオニンの代謝系に大きく影響されるが2),ここでは取り上げない.また,細胞内代謝と密接に関連するmTORシグナルを介したグローバルな翻訳制御についても以下では述べないので,関連する過去の特集号等を参照されたい3).
2)代謝によるRNA結合タンパク質の制御
i) RNAスプライシングの制御
さまざまな代謝関連遺伝子の発現調節において,栄養素や中間代謝産物によるpre-mRNAのスプライシング制御は,重要な役割を果たしている.グルコース-6-リン酸デヒドロゲナーゼ(glucose-6-phosphate dehydrogenase:G6PD)は,解糖系の中間代謝産物であるグルコース6-リン酸をペントースリン酸経路へと流入させる律速酵素であり,近年ではがん代謝における役割が注目されている.G6PDのタンパク質発現は個体の栄養状態に応じ,転写ではなくスプライシングの制御を介して調節されることが知られている4).G6PDのエクソン12にはexonic splicing silencer(ESS)およびexonic splicing enhancer(ESE)が存在しており,絶食状態の肝細胞においてはheterogeneous nuclear ribonucleoprotein(hnRNP)Kの発現が上昇してESSのCリッチ配列へと結合する.それによってserine/arginine-rich splice factor(SRSF)3(下記参照)のESEへの結合が阻害され,スプライシングによるイントロン除去が抑制される.イントロンを含んだままのpre-mRNAは翻訳には使用されずやがて分解されるため,G6PDのタンパク質発現量は低下する5).一方,再摂食後はSRSF3のタンパク質発現およびリン酸化が亢進してエクソン12のESEへ結合し,イントロン除去を促進する.再摂食によるSRSF3のリン酸化にはインスリンシグナルが関与し,アラキドン酸等の不飽和脂肪酸はこれに対して抑制的に働くが6),詳細な分子メカニズムはわかっていない.
SRSFファミリーの機能においてはリン酸化と脱リン酸化のサイクルが重要であることが示唆されているが,上流キナーゼとして知られるClkやSR protein-specific kinase(SRPK)はいずれもAktによって活性化されることから7),代謝によるSRSFを介したスプライシングの制御は,肝細胞のみならずさまざまなコンテクストにおいて起こりうると考えられる.実際に,SRSF3によるG6PDのイントロン除去およびタンパク質発現の促進はHeLa細胞においても確認されており6),上記のメカニズムは種々のがん細胞におけるG6PD発現の制御にも関与していることが示唆される.
種々のビタミンは核内受容体を介して遺伝子の転写制御を行うことがよく知られているが,そのうちいくつかについては,同時にスプライシングも制御することが報告されている.ビタミンAの代謝産物であるレチノイン酸は,レチノイン酸受容体等を介してレチノイン酸応答配列下流の転写を活性化する.このような古典的経路とは異なり,転写制御には必ずしも依存しない,PI3K(phosphoinositide 3-kinase)-AktシグナルおよびMEK(MAPK/ERK kinase)-ERK(extracellular signal-regulated kinase)シグナルの活性化を介した応答が知られている.後者の応答では,レチノイン酸による刺激後,上述のhnRNPファミリーやSRSFファミリーを含むさまざまなスプライシング制御因子群のリン酸化が亢進する8).一方,レチノイン酸による神経系への分化の過程ではSRSF2(別名splicing component 35:SC35)の発現量が亢進し,選択的スプライシングによってPKCδVIII(protein kinase C δVIII)の発現が上昇する9).
ビタミンDの活性型である1,25-ジヒドロキシビタミンD(1,25(OH)2D)についても,スプライシング制御への関与が知られている.1,25(OH)2Dに結合したビタミンD受容体(vitamin D receptor:VDR)は,補助制御因子であるnuclear receptor coactivator(NCoA)/Ski-interacting protein(SKIP)と結合してビタミンD応答配列下流の転写を活性化するが,このときNCoA/SKIPは転写産物のスプライシング制御因子としても機能する10).大腸がん細胞および前立腺がん細胞を用いた解析から,1,25(OH)2Dは自身の代謝酵素であるCYP24の転写とスプライシングを同時に亢進することが確認されている.このとき上記のような転写とスプライシングの共役が起こるか否かは不明であるが,スプライシングの基底活性や1,25(OH)2Dへの応答性が細胞ごとに大きく異なっていることは,非常に興味深い11, 12).
ii) RNA編集の制御
転写後の(pre-)mRNAは,RNA編集(RNA editing)の過程を経ることによって,ゲノムにコードされたものとは一部異なる配列を持つことが可能となる.これまで明らかにされているRNA編集の様式には,特定の塩基が置換される「塩基置換型」と,ウリジンが挿入・欠失する「塩基挿入・欠失型」の二つが存在する.前者は哺乳類を含むさまざまな生物で見つかっており,後者はトリパノソーマのミトコンドリアに特徴的な様式である.塩基置換型のRNA編集は特定の配列で起こり,スプライシングや翻訳に影響するほか,3′非翻訳領域における配列の変換は核内への保持やヌクレアーゼによる分解,マイクロRNAによる標的にも影響する.また,RNA編集は(pre-)mRNAのみならずトランスファーRNAやリボソームRNA,マイクロRNAなどのいわゆる非コードRNAにおいてもみられ,RNA全般の動態を制御する重要なメカニズムの一つとなっている13).
哺乳類でみられる塩基置換型のRNA編集は,ADAR(adenosine deaminase acting on RNA)ファミリーおよびAPOBEC(apolipoprotein B mRNA editing enzyme, catalytic polypeptide-like)ファミリーのデアミナーゼによって行われる13).ADARによるRNA編集はアデノシン(A)からイノシン(I)への編集(A-to-I editing)であり,翻訳の際,イノシンはグアノシンとして認識される.ADARファミリーのうち,ADAR1およびADAR2の標的配列はそれぞれ数千におよび,一部重複する以外はそれぞれ特異的な標的配列が存在している14).ADARファミリーのRNA結合ドメインは二重鎖RNAを認識するが,いずれも配列特異性はみられない15).したがって標的配列の違いは主にデアミナーゼドメインの特異性に依存すると考えられるが,ADAR2では二重鎖RNA結合モチーフによる特異性への寄与が比較的大きいようである16).
興味深いことに,膵β細胞においては,ADAR2によるグルタミン酸受容体サブユニットGluR-BおよびADAR2自身のpre-mRNAの編集は,グルコースやインスリンによって正に制御されている17).このときJNKの活性化を介してADAR2の転写およびタンパク質発現が亢進するが18),より詳細なメカニズムについては解明が待たれる.ADARファミリーの発現変化については,がんを含むさまざまな疾患との相関が見いだされている13, 19).がん細胞のさまざまな代謝変化がADARの発現制御に関与しているか否かは非常に興味深い.
APOBECファミリーはDNAおよびRNAのシチジン(C)からウリジン(U)への編集(C-to-U editing)を行うデアミナーゼであり,さまざまな栄養素や代謝産物,環境ストレス等によってその発現が制御されている.APOBEC1はapolipoprotein B(apoB)のRNA編集を行う酵素として同定され,RNAに対するC-to-U編集についての報告が多くなされている20).一方,APOBEC2, APOBEC3およびAPOBEC4は,相同性に基づくデータベース検索によって同定された.現時点では,APOBEC2についてはデアミナーゼ活性を介する細胞機能の制御が見いだされておらず,APOBEC4については機能解析の報告がない20).APOBEC3はヒトゲノム上に7種類(A, B, C, DE, F, G, H)が存在し,いずれも一本鎖DNAに対する編集活性が知られていたが,最近の研究によりRNAに対する編集活性が見いだされている(下記参照).
肝細胞におけるAPOBEC1の発現は肥満やインスリン,エタノールによって増加することが知られているが,結果としてapoBのRNA編集が亢進して停止コドンが生じ(CAA→UAA),apoB-48の産生が増加する21–23).一方,神経由来の細胞を用いた解析において,低糖濃度・低酸素ストレスによるAPOBEC1の発現亢進が報告されている.このときCOX2(cyclooxygenase 2)のタンパク質発現が亢進するものの,RNA編集は行われず,COX2 mRNAに含まれるAUリッチ配列にAPOBEC1が結合することによってmRNAの安定性を高めていると考えられている24).
APOBEC3のアイソフォームの一つであるAPOBEC3Aは,RNAのC-to-U編集を行うことが近年明らかにされた25).単球からマクロファージへの分化において低酸素シグナルの重要性が示唆されているが,低酸素下の単球ではコハク酸デヒドロゲナーゼBサブユニット(succinate dehydrogenase B:SDHB)のC-to-U編集(R46Xによる不活性化)が著しく促進し,そのほとんどはM1マクロファージにみられること,APOBEC3Aの発現が低酸素環境において亢進し,SDHBを含むさまざまな遺伝子のRNA編集を行うことが示唆されている25).
上記のような栄養素や代謝産物,代謝ストレスによるRNA編集メカニズムへの影響は,担当酵素の発現量やリン酸化の変化を介する例がほとんどであり,それを制御するシグナル経路の活性化を介している.すなわち,あくまで代謝変化による副次的な効果といえる.しかしながら,ゲノムやエピゲノムには必ずしも反映されない多大な影響が代謝変化を背景として生じうることは注目に値する.
3)代謝酵素とRNAの相互作用
さまざまな代謝酵素がDNA・RNAの特定配列に結合して転写や翻訳等の制御に関与することは以前から知られており,いわゆる「moonlighting enzyme(副業を行う酵素)」の代表例となっている26).デヒドロゲナーゼ等にみられるロスマンフォールド(Rossman fold)と呼ばれる高次構造は,代謝反応の補酵素として用いられるNAD+/NADH等のヌクレオチドの結合部位として機能するが,核酸への結合能を併せ持つ例がいくつか知られている.たとえばGAPDHのロスマンフォールドは,mRNAの3′非翻訳領域にみられるAUリッチ配列(AU-rich element:ARE)に結合すること,その結合はNAD+との結合とは競合的に起こることが知られている26).したがって,GAPDHによる代謝反応が盛んに起こる間は,mRNAへの結合は抑制されると考えられる.実際に近年の解析から,T細胞の活性化に伴う酸化的リン酸化から解糖系への代謝スイッチにより,インターフェロンγのmRNAに結合したGAPDHがAREから解離し,翻訳抑制が解除されることが示唆されている27).同様に,乳酸デヒドロゲナーゼ(lactate dehydrogenase:LDH)Aのロスマンフォールドについても,RNAへの結合能が確認されている.LDHAはRNA分解の調節に関与するAUF1(ARE/poly(U)-binding/degradation factor 1)およびHsp(heat shock protein)70と複合体を形成し,RNAへの結合はNAD+との結合と競合的に起こることから,代謝反応とRNA分解制御との共役が示唆される28).
GAPDHと類似の制御機構を示すRNA結合代謝酵素の典型例としては,細胞質型のアコニターゼACO1がよく知られている.ACO1の活性中心である鉄–硫黄クラスターは不安定であり,鉄イオン濃度の減少によってアポ酵素となるが,このときACO1はRNA結合タンパク質として機能する.このような鉄イオンセンサーとしての機能から,ACO1はiron regulatory protein 1(IRP1)としても別途同定されている.ACO1/IRPは,鉄の取り込みを担うトランスフェリンのmRNAの3′非翻訳領域に結合してこれを安定化する一方,鉄貯蔵を行うフェリチンのmRNAの5′非翻訳領域に結合して翻訳を阻害する26).
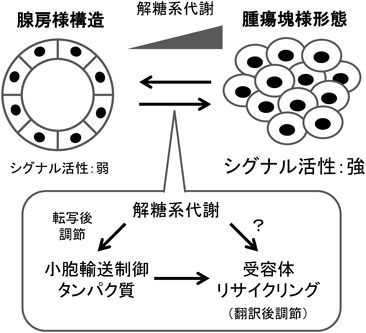
RNA結合タンパク質の網羅的な解析により,さまざまな生化学的コンテクストにおいて,RNA結合能を示す代謝酵素が同定されている.近年moonlighting enzymeとしての役割が特に注目されているピルビン酸キナーゼ(pyruvate kinase:PK)M2など非常に多くの代謝酵素が,上述のロスマンフォールドの有無に関わらず,RNA結合タンパク質として同定されている26, 29).それらの酵素が担う代謝経路は,解糖系,クエン酸回路,核酸合成経路など多岐に及ぶが,上記のACO1/IRP1やGAPDHのような代謝活性と共役した制御の有無については解析が待たれる.興味深いことに,多くのがんで活性が亢進する解糖系については,グルコース6-リン酸の異性化(グルコースリン酸イソメラーゼが行う)とフルクトース6-リン酸のリン酸化(ホスホフルクトキナーゼが行う)の反応を除くすべてのステップにおいて,RNA結合能を持つアイソザイムが一つ以上存在する26, 29).これらにもGAPDHと同様のメカニズムが働くとすれば,がん細胞のワールブルグ効果は,それ自体がRNA動態に非常に大きな影響を与えるものと考えられる.
4)代謝産物の直接的な結合によるRNA動態の制御
主に細菌等のmRNAにみられる「リボスイッチ」と呼ばれる制御配列には,代謝産物などのさまざまな低分子化合物が直接結合して立体構造の変化を誘導し,転写の終結や翻訳の阻害,リボザイム活性による切断等を引き起こすことが知られている30).ピロリン酸チアミン(thiamine pyrophosphate:TPP)が結合するTPPリボスイッチは植物や菌類などの真核生物に広く存在し,pre-mRNAのスプライシング等を制御することが知られているが,動物では見つかっていない.ヒトではVEGFA(vascular endothelial growth factor A)mRNAの3′非翻訳領域にみられる配列が「リボスイッチ」に該当するとされているが,代謝産物ではなくGAIT(interferon-γ-activated inhibitor of translation)およびhnRNP Lなどのタンパク質が相互排他的に結合することで翻訳が調節されており31),上記のようなリボスイッチの定義からは外れている.
一方で,近年の研究から,ATP等のアデノシンやGTP等のグアノシンと結合するRNAアプタマー配列が,ヒトゲノム上にも数多く存在することが明らかにされている32, 33).これらの知見は,リボスイッチのような「代謝産物によるRNA動態の直接的な制御」がヒトの遺伝子発現制御においても広範に行われている可能性を示唆するが,なかでも翻訳やスプライシングの制御に深く関与するグアニン四重鎖(G-quadruplex)構造がGTPへの結合能を持つアプタマーを含むことは,非常に興味深い32).In vitro解析の結果からは,当該配列にはGTPのみならずGDPやGMP, cGMP等も結合しうることが示されているが,生体内における実際の基質特異性や,それぞれの基質が結合した際のRNAへの影響についてはいまだ明らかにされておらず,今後の報告が待たれる.
リボスイッチを介した代謝産物による遺伝子発現の制御は関連する代謝酵素等でみられる場合が多く,ラクトースオペロン等と同様に,常に変動する細胞外栄養素の濃度にすばやく応答するために進化してきた仕組みであると考えられる.このように,代謝産物と被制御遺伝子の関係が明確な場合,リボスイッチの同定にはゲノム解析等のバイオインフォマティクスが非常に有効となる.一方,関連性が容易に想定できない代謝産物と遺伝子との相互作用の同定には生化学的な手法に基づく網羅的な解析が必要である.上記のグアノシン結合配列はそのような解析によって同定された例であるが,今後の解析手法のさらなる発展によって,代謝産物そのものによるRNA制御の仕組みが明らかになるかもしれない.
引用文献References
1) Castello, A., Hentze, M.W., & Preiss, T. (2015) Trends Endocrinol. Metab., 26, 746–757.
2) Maddocks, O.D., Labuschagne, C.F., Adams, P.D., & Vousden, K.H. (2016) Mol. Cell, 61, 210–221.
3) 高原照直,前田達哉(2013)生化学,85, 205–213.
4) Salati, L.M., Szeszel-Fedorowicz, W., Tao, H., Gibson, M.A., Amir-Ahmady, B., Stabile, L.P., & Hodge, D.L. (2004) J. Nutr., 134, 2437S–2443S.
5) Cyphert, T.J., Suchanek, A.L., Griffith, B.N., & Salati, L.M. (2013) Biochim. Biophys. Acta, 1829, 905–915.
6) Walsh, C.M., Suchanek, A.L., Cyphert, T.J., Kohan, A.B., Szeszel-Fedorowicz, W., & Salati, L.M. (2013) J. Biol. Chem., 288, 2816–2828.
7) Zhou, Z. & Fu, X.D. (2013) Chromosoma, 122, 191–207.
8) Laserna, E.J., Valero, M.L., Sanz, L., del Pino, M.M., Calvete, J.J., & Barettino, D. (2009) Mol. Endocrinol., 23, 1799–1814.
9) Apostolatos, H., Apostolatos, A., Vickers, T., Watson, J.E., Song, S., Vale, F., Cooper, D.R., Sanchez-Ramos, J., & Patel, N.A. (2010) J. Biol. Chem., 285, 25987–25995.
10) Zhang, C., Dowd, D.R., Staal, A., Gu, C., Lian, J.B., van Wijnen, A.J., Stein, G.S., & MacDonald, P.N. (2003) J. Biol. Chem., 278, 35325–35336.
11) Muindi, J.R., Nganga, A., Engler, K.L., Coignet, L.J., Johnson, C.S., & Trump, D.L. (2007) J. Steroid Biochem. Mol. Biol., 103, 334–337.
12) Peng, X., Tiwari, N., Roy, S., Yuan, L., Murillo, G., Mehta, R.R., Benya, R.V., & Mehta, R.G. (2012) J. Endocrinol., 212, 207–215.
13) Avesson, L. & Barry, G. (2014) Biochim. Biophys. Acta, 1845, 308–316.
14) Wang, I.X., So, E., Devlin, J.L., Zhao, Y., Wu, M., & Cheung, V.G. (2013) Cell Reports, 5, 849–860.
15) Nishikura, K., Yoo, C., Kim, U., Murray, J.M., Estes, P.A., Cash, F.E., & Liebhaber, S.A. (1991) EMBO J., 10, 3523–3532.
16) Eggington, J.M., Greene, T., & Bass, B.L. (2011) Nat. Commun., 2, 319.
17) Gan, Z., Zhao, L., Yang, L., Huang, P., Zhao, F., Li, W., & Liu, Y. (2006) J. Biol. Chem., 281, 33386–33394.
18) Yang, L., Huang, P., Li, F., Zhao, L., Zhang, Y., Li, S., Gan, Z., Lin, A., Li, W., & Liu, Y. (2012) PLoS ONE, 7, e48611.
19) Zipeto, M.A., Jiang, Q., Melese, E., & Jamieson, C.H. (2015) Trends Mol. Med., 21, 549–559.
20) Smith, H.C., Bennett, R.P., Kizilyer, A., McDougall, W.M., & Prohaska, K.M. (2012) Semin. Cell Dev. Biol., 23, 258–268.
21) Phung, T.L., Sowden, M.P., Sparks, J.D., Sparks, C.E., & Smith, H.C. (1996) Metabolism, 45, 1056–1058.
22) von Wronski, M.A., Hirano, K.I., Cagen, L.M., Wilcox, H.G., Raghow, R., Thorngate, F.E., Heimberg, M., Davidson, N.O., & Elam, M.B. (1998) Metabolism, 47, 869–873.
23) Lau, P.P., Cahill, D.J., Zhu, H.J., & Chan, L. (1995) J. Lipid Res., 36, 2069–2078.
24) Li, W., Cheng, X., Chen, H.S., & He, Z.Y. (2013) Neurochem. Res., 38, 1434–1445.
25) Sharma, S., Patnaik, S.K., Taggart, R.T., Kannisto, E.D., Enriquez, S.M., Gollnick, P., & Baysal, B.E. (2015) Nat. Commun., 6, 6881.
26) Castello, A., Hentze, M.W., & Preiss, T. (2015) Trends Endocrinol. Metab., 26, 746–757.
27) Chang, C.H., Curtis, J.D., Maggi, L.B. Jr., Faubert, B., Villarino, A.V., O’Sullivan, D., Huang, S.C., van der Windt, G.J., Blagih, J., Qiu, J., Weber, J.D., Pearce, E.J., Jones, R.G., & Pearce, E.L. (2013) Cell, 153, 1239–1251.
28) Pioli, P.A., Hamilton, B.J., Connolly, J.E., Brewer, G., & Rigby, W.F. (2002) J. Biol. Chem., 277, 35738–35745.
29) Castello, A., Fischer, B., Eichelbaum, K., Horos, R., Beckmann, B.M., Strein, C., Davey, N.E., Humphreys, D.T., Preiss, T., Steinmetz, L.M., Krijgsveld, J., & Hentze, M.W. (2012) Cell, 149, 1393–1406.
30) Serganov, A. & Nudler, E. (2013) Cell, 152, 17–24.
31) Ray, P.S., Jia, J., Yao, P., Majumder, M., Hatzoglou, M., & Fox, P.L. (2009) Nature, 457, 915–919.
32) Vu, M.M., Jameson, N.E., Masuda, S.J., Lin, D., Larralde-Ridaura, R., & Lupták, A. (2012) Chem. Biol., 19, 1247–1254.
33) Curtis, E.A. & Liu, D.R. (2013) Chem. Biol., 20, 521–532.
34) Emerman, J.T., Bartley, J.C., & Bissell, M.J. (1981) Exp. Cell Res., 134, 241–250.
35) Onodera, Y., Nam, J.M., & Bissell, M.J. (2014) J. Clin. Invest., 124, 367–384.
36) Onodera, Y., Nam, J.M., & Sabe, H. (2013) Pharmacol. Ther., 140, 1–9.
37) Kao, A.W., Noda, Y., Johnson, J.H., Pessin, J.E., & Saltiel, A.R. (1999) J. Biol. Chem., 274, 17742–17747.
38) Tisdale, E.J., Kelly, C., & Artalejo, C.R. (2004) J. Biol. Chem., 279, 54046–54052.
39) Zala, D., Hinckelmann, M.V., Yu, H., Lyra da Cunha, M.M., Liot, G., Cordelières, F.P., Marco, S., & Saudou, F. (2013) Cell, 152, 479–491.
40) Podbilewicz, B. & Mellman, I. (1990) EMBO J., 9, 3477–3487.