ABO血液型や,腫瘍マーカーとして臨床で用いられている抗原の多くが糖鎖であることはよく知られている.たとえば,消化器がんで陽性になることが多いCA19-9はシアリルLewis A(Siaα2-3Galβ1-3[Fucα1-4]GlcNAcβ1-3Gal)が抗原であり,SLX(シアリルLewis X, Siaα2-3Galβ1-4[Fucα1-3]GlcNAcβ1-3Gal)は肺がんや消化器がんなどで検出される.糖鎖が細胞表面に発現するため,マーカーとして利用しやすいことや,がん化や悪性度の上昇によって糖転移酵素遺伝子の発現パターンがしばしば大きく変わること,それによって糖鎖パターンも劇的に変わりうること,などの理由で糖鎖が腫瘍マーカーとして多用されていると考えられる.加えて,がん細胞に特有なさまざまなタイプの糖鎖構造は,抗がん剤などの特異的なターゲットとなる可能性があり,治療への応用も期待されている.
マウス胚性幹細胞(ES細胞)の未分化性を確認するマーカーとしてよく用いられるSSEA-1は,F9胚性がん腫細胞をBALB/cマウスに免疫して得られたモノクローナル抗体が認識する抗原1)で,その一部分には抗原エピトープとしてLewis X(Lex,Galβ1-4[Fucα1-3]GlcNAc)を含む2).SSEA-1とLexは厳密には同じものではないが,よく同じものとして扱われ,文献的にも完全に区別されているわけではない.正常発生では8細胞期~桑実胚期の胚や,胚盤胞の内部細胞塊から得られるES細胞,あるいはiPS細胞など,多能性を示す幹細胞で豊富に発現している.マウスES細胞におけるSSEA-1の発現は,分化条件で培養することにより,他の未分化マーカー(たとえばTra1-60,これも糖鎖抗原である)とともに速やかに消失することから,未分化性の維持に何らかの機能を有すると考えられてきたが,その仮説はいまだ証明されていない.さらに,仮説が正しいとして,その分子機序もよくわかっていない.ちなみに,ヒトES細胞ではSSEA-1は未分化マーカーとしては不適で,SSEA-3/4(これもグロボ系列糖脂質の糖鎖抗原)などが未分化マーカーとして用いられることから,SSEA-1抗原発現と未分化性との機能的関連が疑問視されることもある.しかしながら,当研究室のHPLCシステム(後述)を用いた詳細な解析では,ヒトES細胞にもLexが発現していることを確認した(未発表データ).量は少なくとも,未分化性の維持に必須の糖タンパク質/糖脂質の上にのみ,Lexが発現していれば用は足りるのかもしれないのである.
その後,SSEA-1もしくはLexを認識する抗体(たとえば,CD15など)によって組織幹細胞の一部が染色されることが報告された.特に,マウスやヒトの胎生期および成体脳の神経幹細胞に,Lexが豊富に発現することが示され,SSEA-1/Lexの発現はES細胞以外の幹細胞でも未分化性を示すと考えられるようになった3).細胞表面に発現するLexに対する抗体と,蛍光フローサイトメーター(FACS)を用いることにより,容易にLex発現細胞を分離・回収することが可能だが,脳室周囲組織から分離したLex発現細胞には,神経幹細胞が高密度に存在することが報告された4).神経幹細胞や網膜幹細胞は,線維芽細胞増殖因子2(FGF2)や上皮増殖因子(EGF)を添加した無血清培地で培養することができ,neurosphereと呼ばれる浮遊細胞塊を形成する4, 5).Lexに対するFACSによって,成体脳の神経幹細胞のほか,マウス胎仔脳の神経幹細胞や,マウス胎仔網膜から網膜幹細胞などが濃縮され,より多くのneurosphereが形成されることが報告されている3).
本稿では,未分化性の維持に重要なLexの生合成に関与するα1,3-フコース転移酵素Fut9およびFut10の神経幹細胞での役割について述べる.
2. 脳のLewis Xとα1,3-フコース転移酵素
Lexの生合成は,N-アセチルラクトサミンにα1,3結合でフコースを加える反応が律速であり,α1,3-フコース転移酵素によって触媒される.げっ歯類には3種類の活性型α1,3-フコース転移酵素(Fut4, Fut7, Fut9)が存在するが,脳内の糖タンパク質・糖脂質のLexのほとんどはFut9が合成すると考えられていた.未分化な細胞に特異的に発現しているからといって,未分化性に機能的に寄与しているかどうかはわからないが,神経幹細胞に発現するLexを生合成するFut9は,神経発生に重要な働きを持つと予想されていた.しかしながら,Fut9ノックアウトマウスでは脳構造に大きな異常は観察されず,通常どおりに発生・出生・発育して,不安様行動の増加以外はおおむね正常な行動を示した6).
我々は,神経幹細胞の未分化性維持におけるFut9の役割を調べるため,Fut9ノックアウトマウス胎仔脳から,neurosphere法で神経幹細胞の培養を行った7).形成されたneurosphereを再度分散し,同様の条件で培養することにより自己複製能(ここでは未分化性維持能力とほぼ同義である)を,接着させて分化条件で培養することにより神経細胞・アストロサイト・オリゴデンドロサイトの3種類の細胞へ分化できる多分化能を計測することができる.その結果,Fut9−/−胎仔脳から培養した神経幹細胞は,予想に反してより高い自己複製能を示すことがわかり,Fut9もしくはそれが生合成するLexは,むしろ神経幹細胞の分化を促進していることが示唆された7).このことから,Lexが神経幹細胞の未分化性の維持と自己複製能に重要であるとすると,Fut9以外のα1,3フコース転移酵素によってLexの合成が行われている可能性が示唆された.
3. 幹細胞の未分化性維持に関わるα1,3-フコース転移酵素
2000年前後に次々に解読されたヒトやマウスなどの全ゲノム情報を基に,Fut9までの9種類のFut遺伝子に加え,さらに,2種類の遺伝子Fut10とFut11が,アミノ酸配列上他のα1,3-フコース転移酵素遺伝子群とファミリーを形成することが報告された8).これらの遺伝子は脳で豊富に発現するものの,培養細胞での過剰発現や,通常のα1,3-フコース転移酵素活性測定では,Lex合成活性が検出されなかった9).その後,Fut11がGDP-フコースの加水分解活性を持つこと10)や,Fut10がN-グリカンの内側のN-アセチルグルコサミンにフコースを転移すること11)が報告されたが,これらの酵素がLexを合成できるα1,3-フコース転移活性を持つかどうかは,依然として不明であった.
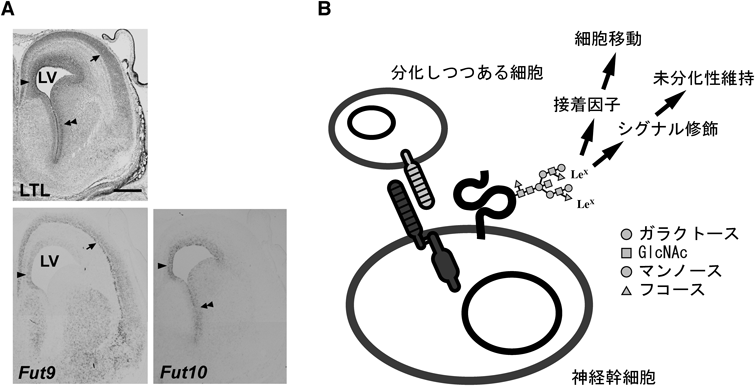
我々は,Fut10とFut11が神経幹細胞におけるLex発現に関わっているかどうかを調べるため,マウス胎生15.5日胚の脳におけるそれらの発現部位を,in situハイブリダイゼーション法で検索した.その結果,神経幹細胞が存在する脳室帯にFut10が豊富に発現していることが判明した7)(図1A).Fut9の発現は脳室帯にも認められるものの,背内側に限局しており,神経幹細胞が多数存在する腹外側の基底核原基(ganglionic eminence: GE)には検出できなかった.むしろ,神経幹細胞が分化して放射状移動していく皮質板にFut9の発現が強く観察され,Fut9が神経幹細胞の分化と関係していることを示唆する上記の結果と一致した.そこで我々は,背側と腹側の大脳を区別して糖鎖解析することにより,Fut10のα1,3-フコース転移酵素活性について何らかの知見が得られることを期待した.
生理学研究所・池中研究室では,イオン交換カラムに加えて順相・逆相の3種類のHPLCを組み合わせることにより,糖タンパク質N結合型糖鎖の網羅的解析を行っている(3D HPLC法).さらに,ヒドラジン分解や還元末端のピリジルアミノ化などの処理をカラム内で行うことによって微量サンプルの解析法を確立しており,マウス脳などに存在するN-グリカン糖鎖のマッピングをすでに終えている12).この3D HPLC技術でマウス脳を詳細に解析したところ,Fut9が発現する背側の大脳皮質には,バイアンテナ型のLex含有糖鎖が認められたのに対し,Fut9が発現しない(ただしFut10は存在する)GEの脳室帯には,分岐型のLex含有糖鎖が検出された7).このことから,Lex含有糖鎖構造の違いによって,機能が異なり,Fut9が発現しない領域で神経幹細胞が多数存在することから,未分化性の維持にはFut10を含む他のα1,3-フコース転移酵素活性を持つ酵素が関与していると考えられる.
一方,in vitro α1,3-フコース転移酵素活性測定では,分岐型N-グリカンを含む各種の糖鎖基質を試したにも関わらず,Fut10のα1,3-フコース転移酵素活性は検出できなかった.非還元末端にGalβ1-4GlcNAcを含む糖鎖であれば基質にできるFut9と異なり,Fut10は(もしかするとペプチド部分も含む)非常に厳密な基質特異性を持つ可能性が考えられる.そのため,今後Fut10がどのような糖鎖もしくは糖タンパク質を基質にできるのか調べる必要がある.
Fut10に対するmiRNA発現ベクターを用い,ES細胞でノックダウン実験を行った7).マウスのES細胞は,以前より使用されているLIF(leukaemia inhibitory factor)に加え2種類の阻害剤(MEK阻害剤+GSK3阻害剤)存在下では,未分化な状態で安定して培養することができ13),Oct3/4やNanogのような未分化マーカーを高発現する.このような培養条件でFut10をノックダウンしたところ,Oct3/4,Nanogの発現が著明に減少し,神経系の分化マーカーであるSox2や,初期内胚葉マーカーであるGata4の発現が上昇した.
また,前述したマウス胎仔脳由来神経幹細胞の培養系を用い,レトロウイルスなどを用いてFut10の過剰発現/ノックダウンを行ったところ,Fut10過剰発現で神経幹細胞の自己複製能が上昇し,ノックダウンでは逆の結果が得られた.これらの結果は,野生型神経幹細胞のみならず,Fut9−/−神経幹細胞でも同様に観察されたことから,Fut10が(神経)幹細胞の自己複製能を亢進させ,その働きはFut9に依存していないことが示唆された.
生体内での神経発生におけるFut10の役割を解明するため,子宮内電気穿孔法を用いて,Fut10のノックダウン実験を行った7).胎生期の脳では,脳室帯に存在する神経幹細胞は,増殖を終えて分化し始めると,放射状に皮質板へと移動する.その際,脳室帯マーカーであるPax6の発現を落とし,脳室下帯マーカーであるTbr2を発現するようになる.胎生14.5日マウス胚の脳で,Fut10をノックダウンしたところ,BrdU取り込みが低下し,Pax6陽性細胞が減少してTbr2陽性細胞が増加することがわかった.これらの結果は,Fut10によって,未分化な状態に維持されていた神経幹細胞が,Fut10ノックダウンによって分化する方向へ移行したことを示しており,上記の結果と一致する.一方,胎生14.5日脳でFut10をノックダウンして,3日後に細胞の分布を検討したところ,放射状移動が著明に抑制されていることが判明した.この結果は,Fut10ノックダウンによる神経幹細胞の分化促進では説明できない(むしろ逆)ので,Fut10は神経幹細胞の分化(未分化性の維持)と移動を独立に制御している可能性が考えられた(図1B).
これまでの研究から,Fut10は非常に厳密な基質特異性を持つα1,3-フコース転移酵素だと考えられた.Fut9とFut10とでは,Lex糖鎖を付加するN結合型糖鎖(もしくは糖タンパク質そのもの)が異なることで,異なるLex含有糖鎖の機能を発揮することが示唆された.すなわち,Fut9は神経幹細胞の分化を促進するのに対し,Fut10は逆に神経幹細胞の未分化性維持に重要であると考えられる.しかしながら,生体内において,神経幹細胞が分化するときに,どのようにFut10の発現の調節もしくは糖鎖構造の変化が起き,分化を促進するFut9の発現,糖鎖構造の変化が起きるのかはいまだに明らかでない.Lex含有糖鎖の違いによって,どのように細胞の性質が変化するのかを明らかにすることで,Fut9とFut10の役割の違いもみえてくると考えられる.
幹細胞の未分化性維持におけるLexの機能,あるいはそれを生合成するFut10の役割が,おぼろげながらみえてきたように思われる.しかしながら,今回の研究では手つかずのFut11の役割や,どのようなキャリアタンパク質に付加されたLexが機能的に重要なのかなど,未解明の問題が山積されている.神経(幹)細胞上に発現している,Lexを含む糖タンパク質については,最近のみにれびゅうでも詳述されている14)ので,参照されたい.
謝辞Acknowledgments
Fut9ノックアウトマウスの解析は産業技術総合研究所糖鎖医工学研究センター・成松久センター長との共同研究で,N結合型糖鎖の網羅的解析は生理学研究所分子神経生理部門・池中一裕教授との共同研究です.池中研究室の皆さんには大変お世話になり,深く感謝しております.
引用文献References
1) Solter, D. & Knowles, B.B. (1978) Proc. Natl. Acad. Sci. USA, 75, 5565–5569.
2) Gooi, H.C., Feizi, T., Kapadia, A., Knowles, B.B., Solter, D., & Evans, M.J. (1981) Nature, 292, 156–158.
3) Yanagisawa, M. & Yu, R.K. (2007) Glycobiology, 17, 57R–74R.
4) Capela, A. & Temple, S. (2002) Neuron, 35, 865–875.
5) Reynolds, B.A. & Weiss, S. (1992) Science, 255, 1707–1710.
6) Kudo, T., Fujii, T., Ikegami, S., Inokuchi, K., Takayama, Y., Ikehara, Y., Nishihara, S., Togayachi, A., Takahashi, S., Tachibana, K., Yuasa, S., & Narimatsu, H. (2007) Glycobiology, 17, 1–9.
7) Kumar, A., Torii, T., Ishino, Y., Muraoka, D., Yoshimura, T., Togayachi, A., Narimatsu, H., Ikenaka, K., & Hitoshi, S. (2013) J. Biol. Chem., 288, 28859–28868.
8) Roos, C., Kolmer, M., Mattila, P., & Renkonen, R. (2002) J. Biol. Chem., 277, 3168–3175.
9) Patnaik, S. (2007) Nature Precedings ‹http://hdl.handle.net/10101/npre.2007.141.1›
10) Baboval, T. & Smith, F.I. (2002) Mamm. Genome, 13, 538–541.
11) Mollicone, R., Moore, S.E., Bovin, N., Garcia-Rosasco, M., Candelier, J.J., Martinez-Duncker, I., & Oriol, R. (2009) J. Biol. Chem., 284, 4723–4738.
12) Yoshimura, T., Yamada, G., Narumi, M., Koike, T., Ishii, A., Sela, I., Mitrani-Rosenbaum, S., & Ikenaka, K. (2012) Anal. Biochem., 423, 253–260.
13) Ying, Q.L., Wray, J., Nichols, J., Batlle-Morera, L., Doble, B., Woodgett, J., Cohen, P., & Smith, A. (2008) Nature, 453, 519–523.
14) 矢木宏和,加藤晃一(2013)生化学,85, 1012–1016.
著者紹介Author Profile
 鈴木 春満(すずき はるみつ)
鈴木 春満(すずき はるみつ)滋賀医科大学医学部生理学講座統合臓器生理学部門博士研究員.博士(バイオサイエンス).
略歴2007年東京工科大学バイオニクス学部(現:応用生物学部)卒業,09年京都大学大学院理学研究科生物科学専攻卒業,14年奈良先端科学技術大学院大学バイオサイエンス研究科神経機能科学研究室卒業.15年より,現職.
研究テーマと抱負神経幹細胞の未分化能の維持に重要なα1,3フコース転移酵素Fut10の働きや糖鎖構造を変化させることで分化の制御ができないかと野望を抱いている.
ウェブサイトhttp://www.shiga-med.ac.jp/~hqphysi1/(研究室)
趣味伝統的なお祭りや神社・お寺などを訪れて写真を撮ること.地元グルメの食べ歩き.