生体を構成するすべての細胞は,重力,引張力など多様な力学的環境にさらされている.細胞は,細胞内外の機械的シグナル(メカニカルストレス)を感知し,これを化学的シグナルに変換して,その形態や増殖・分化・運動能を変化させる.このように細胞が機械的シグナルを感知・伝達し,応答する機能は細胞の力覚応答と呼ばれ,生体の発生過程や恒常性維持において重要な役割を担っている.その破綻は器官形成不全,筋萎縮,骨粗鬆症,循環器疾患,がんなど多様な疾患の発症にも関与しており,生理的・病理的にも重要である.
細胞は,力学的シグナルに応答して細胞骨格を再構築する.たとえば,血管内皮細胞は,血流や血圧による剪断応力や伸展張力などの力学的シグナルによって細胞骨格と細胞形態を変化させ,血管内環境の恒常性維持に寄与している1).また,上皮細胞は,細胞外基質の硬さに依存して細胞骨格や細胞接着を変化させ,運動能や増殖・分化・細胞死などを制御することもよく知られている2).機械的シグナルを受容・伝達するメカノセンサー分子としては,以前から細胞膜上の機械受容チャネルが同定されており,細菌の機械受容チャネルについては機械的刺激による立体構造変化など詳しい解析がなされている3, 4).一方,近年,非チャネル型の新たなメカノセンサー分子として,細胞骨格や細胞接着に関わる分子が次々と同定されており,細胞の力覚応答における細胞骨格,細胞接着の重要性が注目されている.本稿では,細胞の力覚応答における細胞骨格,細胞接着装置の役割と,力学的刺激による細胞骨格再構築の制御機構について概説する.
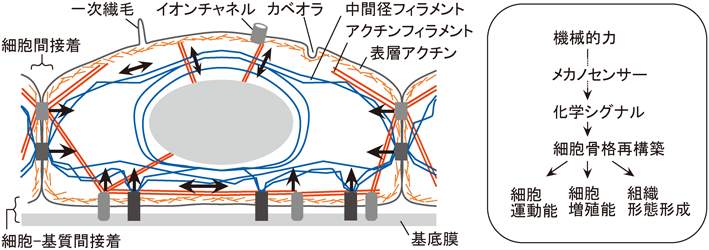
細胞骨格はアクチン繊維,微小管,中間径フィラメントから構成される.アクチン繊維と中間径フィラメントは細胞間や細胞–基質間接着装置に繋留されており,細胞全体に網目状のネットワークを構築している.したがって,これらの細胞骨格や細胞接着装置は,細胞に外力が加わった際に最初に力を感知するメカノセンサー装置として機能すると考えられる(図1).一方,各々の細胞骨格は異なる性質を持ち,力覚応答における機能も異なることが明らかとなってきた.本節では各々の細胞骨格のメカノセンサーとしての機能について述べる.
1)アクチン繊維とメカノセンシング
アクチン繊維は単量体アクチンの重合により形成される.アクチン繊維は動的な性質を持ち,細胞内では多くの結合タンパク質によって,重合・脱重合や束化や分枝・網目構造の形成と崩壊が制御されている.このようなアクチン骨格の再構築は細胞の形態変化,運動,接着,極性化,分裂,輸送など多様な細胞活動において中心的な役割を担っている.細胞に機械的刺激が加わると,一過的にアクチン繊維が崩壊し単量体アクチン濃度が増加する.このとき,アクチン結合タンパク質フォルミンファミリーが活性化されアクチン重合核を形成し,新たなアクチン繊維の形成が促進される5).また,本稿では詳しく述べないが,機械的刺激によるアクチン重合は転写制御因子の核移行を促進し,細胞の増殖や分化を制御している.転写因子SRFの活性化因子であるmegakaryoblastic leukemia (MKL)/myocardin-related transcription factor (MRTF)は,通常の条件ではアクチン単量体と結合して細胞質にとどまっているが,血清刺激や機械的刺激によってアクチン重合が促進されるとアクチン単量体から解離し,核移行と標的遺伝子の転写が促進される6, 7).また,Hippo経路によって負に制御される転写制御因子であるYAP/TAZも,機械的刺激によってアクチン重合が促進されると,核移行が促進される8).このように,アクチン骨格はメカニカルストレスにより迅速に再構築される構造であり,力覚応答における役割やその分子機構が注目されている.
ストレスファイバーは非筋細胞で観察される10~30本のアクチン繊維とミオシンからなる繊維束で,細胞に収縮力を発生させる主要な構造である.ストレスファイバーを構成するアクチン繊維はα-アクチニンによって架橋されており,非筋型ミオシンとトロポミオシンが周期的に分布している9).ストレスファイバーは両端が細胞–基質間接着装置である接着斑(focal adhesion)や細胞間接着装置であるアドヘレンスジャンクション(AJ)に接続されており,接着端間に収縮力を発生させる.ストレスファイバーは古くから細胞機能に重要な役割を果たすと考えられ,その性質が研究されてきた10, 11).近年,アクチン繊維自体がメカノセンサーとして機能することが示された.単離したストレスファイバーを用いて,アクチン繊維は引張された状態ではアクチン切断因子であるコフィリンが結合しにくくなり,コフィリンによる切断を受けにくいことが示された.さらに,血管内皮細胞を用いて,コフィリンは弛緩した状態のストレスファイバーに結合しやすいことが示された12).アクチン繊維は右巻きらせん構造をとるが,コフィリンが結合したアクチン繊維ではらせんがさらに右にねじれた構造を持つことが電子顕微鏡解析により示されている13).アクチン繊維は引張によってらせんのピッチが長くなるとコフィリンが結合しにくい構造となり安定化され,弛緩してピッチが短くなるとコフィリンが結合しやすくなり切断されやすくなると考えられる.このように,アクチン繊維は引張力の負荷・消失でそれ自身の安定性が制御されるメカノセンサーとしての機能を持つと考えられる.
また,アクチン繊維の伸長はアクチン繊維とミオシンIIの相互作用を促進することも示されている14).蛍光タンパク質GFPを付加したミオシンIIのサブフラグメント(S1)を細胞性粘菌に発現させ,これをプローブとして,GFP-S1がより張力のかかる部位にあるアクチン繊維と結合することが示された.アクチン繊維のピッチの長さは,ミオシンが結合した状態では0.2%長いことがX線構造解析により示されており15),ミオシンIIは伸長されたアクチン繊維に対してより高い親和性を持つと考えられる.引張力によってアクチン繊維が伸長するとミオシンIIの結合が促進され収縮力が発生すると推測され,この場合もアクチン繊維自身がメカノセンサーとして力学的刺激に応答して,フィードバック的に機能している可能性が考えられる.これらの報告から,アクチン繊維には,らせん構造の伸縮性により,引張される部位のストレスファイバーではミオシンが結合し張力を発生し,逆に,張力のかからない部位ではコフィリンが結合し,不要なストレスファイバーが崩壊するという合理的なシステムが存在することが推定される16).アクチン繊維は筋細胞,非筋細胞いずれにおいても,コフィリンやミオシン以外にも多様な結合タンパク質による制御を受けている.引張によるアクチン繊維の構造変化はこれらの他のアクチン結合タンパク質の結合・解離にも影響する可能性があり,今後の研究の進展が期待される.
2)微小管とメカノセンシング
微小管はアクチン繊維と同様にすべての真核生物が保有する細胞骨格であり,細胞内輸送,分裂,繊毛形成など,多様な細胞機能に関与している.微小管はαチューブリンとβチューブリンが1 : 1で結合したヘテロ二量体が重合したプロトフィラメントを基本構造とし,これが13本結合した管状の構造をとる.微小管は,中心体や紡錘体極など微小管形成中心を重合核として主に形成され,重合・脱重合で伸長と崩壊が起こる動的不安定性を持つ繊維構造であり,種々の結合因子により重合・脱重合の制御を受ける他,微小管自体の翻訳後修飾によりその安定性が制御されている17, 18).
微小管は,浸透圧や力の負荷によって,その長さが変化することや,張力の負荷で形成が促進されることが報告されており,メカノセンシングとの関連が示唆されている19, 20).しかし微小管自体にメカノセンサー機能があるかどうかは不明である.線虫Caenorhabditis elegansのβチューブリンをコードするmec-7の欠失変異体は触覚を欠く変異体であるが,触覚ニューロンにおける接触依存的な受容体電位変化は,mec-7の欠損で減少するものの,完全には喪失しない21).このことから,少なくともこの系では力を受容する過程に微小管は必須ではないと考えられる22).
3)中間径フィラメントとメカノセンシング
中間径フィラメントは多くの動物細胞に存在する丈夫な繊維構造であり,核を取り囲む密な網目構造が辺縁部まで到達し,細胞間接着部位ではデスモソームに,細胞–基質間接着部位ではヘミデスモソームに繋留されている.中間径フィラメントの構成タンパク質はタイプI~Vに分類され,組織特異的な発現分布を示す.基本的な構造は共通しており,ヘッド,ロッド,テイルの三つのドメインからなり,ロッドドメインで会合して二量体を作り,これが結合して基本構造となる四量体を形成し,この四量体がさらに重合して中間径フィラメントとなる.たとえば,上皮細胞で特異的に発現するケラチン繊維はタイプIとタイプIIが会合したヘテロ二量体から形成され,また間葉系細胞や未分化の細胞に存在するビメンチンはタイプIIIに属し,1種類のタンパク質のホモ二量体から形成される.中間径フィラメントは不溶性が高く,当初は安定で静的な構造と考えられていたが,重合と脱重合を繰り返す動的な構造であることや,リン酸化で速やかに脱重合するなど種々の翻訳後修飾により重合・脱重合や安定性が制御されていることが明らかとなり23, 24),現在では動的に構造変化する細胞骨格であると考えられている.
中間径フィラメントは上皮細胞,筋細胞,ニューロン軸索など,力学的負荷が強くかかる細胞に多く発現しており,細胞に構造的な強度を与え,力学的負荷に対して細胞構造を保守する働きを持つ25, 26).単離した中間径フィラメントは平均2.6倍も伸張することや27),細胞への力負荷によって中間径フィラメントが変形することも生細胞で観察されており28),メカノセンシングへの関与も示唆されている.中間径フィラメントのうちビメンチンは,細胞の収縮力の有無で構造変化することが示されている.システインショットガン法(タンパク質が構造変化した際に,タンパク質の内部から表面に露出するシステイン残基を検出する方法)により,ミオシンII阻害剤Blebbistatinの有無で構造変化する細胞内タンパク質としてビメンチンが同定された29).また,後述のように,我々は,単層上皮細胞で主に発現するケラチン18の発現を抑制すると,力学的刺激で誘発されるRhoAの活性化やストレスファイバーの形成が抑制されることを明らかにした30).このように,メカノセンシングにおいて中間径フィラメントは重要な役割を担っていると考えられるが,中間径フィラメント自身がメカノセンサーとして機能しているかどうかなど,その分子機構は不明であり,今後解決すべき課題である.
上皮細胞が隣接細胞間に形成する細胞間接着装置や,細胞外基質との間に形成する細胞–基質間接着装置にはアクチン繊維や中間径フィラメントが繋留されている(図1).MDCK細胞では細胞間におよそ100 nNの力が発生し,互いに張力を伝えている31).力学的シグナルは主に細胞間や細胞–基質間接着部位から入力され,その結果起こるシグナル伝達,細胞応答によって接着部位の接着複合体や接着部位に繋留される細胞骨格が増強される32).実際にアクチン繊維や微小管を蛍光タンパク質で可視化した細胞を用いて,ガラス微細針で局所を引っぱると,引っぱった方向と平行にストレスファイバーや微小管が形成,増強される20).細胞がいかにして接着部位で力を感知し,化学的シグナルに変換し,細胞骨格を増強するのか,本節では上皮細胞の接着部位におけるメカノセンシング機構や,力学的刺激による接着構造の増強機構について,概説する.
1)アドヘレンスジャンクション(AJ)とメカノセンシング
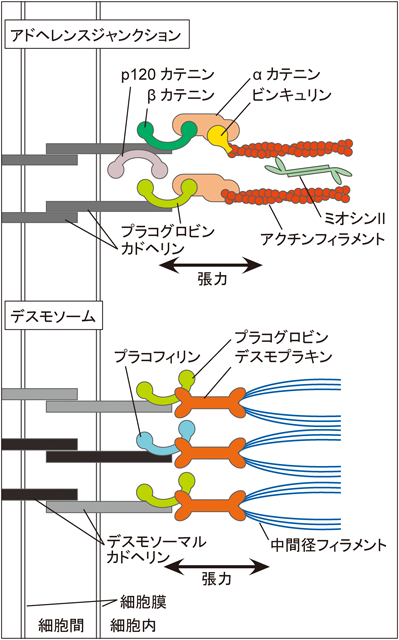
AJは隣接細胞間の接着部位において,膜貫通タンパク質であるカドヘリンが細胞外で同種結合し,細胞内ではαカテニン,βカテニン,ビンキュリンなどと複合体を形成し,アクチン繊維を繋留する接着構造である(図2上).上皮細胞ではAJは細胞の頂端側で,タイトジャンクション(TJ)よりも基底側に形成され,細胞から細胞へと連続して形成され接着帯を構築する.この性質によって上皮細胞シートの集団としての運動や形態形成が可能となる.AJの成熟にはアクチン骨格依存的な張力が重要であることが,AJに発生する張力を測定する実験によって示されている33).この現象はカドヘリンをコートしたビーズを細胞に付着させ,細胞間の張力を模した引っぱり刺激を与えるという,より直接的な手法でも証明されている34).また,張力のかかる細胞間接着部位にはビンキュリンが集積するが34),このとき,αカテニンがメカノセンサーとしてビンキュリンの集積に関与することが示されている35).αカテニンの種々の欠失変異体を用いた実験によって,αカテニンのビンキュリン結合部位は通常は分子内に隠れた状態にあるが,カドヘリン–αカテニン複合体にストレスファイバーからの張力が加わると,張力依存的にαカテニンの中央ドメインが引き伸ばされてビンキュリン結合部位が露出し,ビンキュリンが結合できるようになり,ビンキュリンの集積とAJの成熟が起こるというモデルが提唱されている.
2)デスモソームとメカノセンシング
上皮細胞では,デスモソームが細胞間接着部位に形成される.デスモソームではAJとは異なる種類のカドヘリン(デスモソーマルカドヘリン)が細胞外で同種結合し,細胞内ではプラコフィリン,プラコグロビン,デスモプラキンなどと複合体を形成し,そこにケラチン繊維が繋留されている(図2下).未分化の上皮細胞のデスモソームはAJと同様にカルシウム依存的だが,分化に応じてカルシウム非依存的なより安定な接着構造を形成し,これが上皮組織のバリア機能やメカニカルストレスを受けた際の構造維持などの耐性に重要であることが知られている.近年,張力依存的にデスモソームが増強されることが報告され,メカノセンシングにおけるデスモソームと中間径フィラメントの重要性が注目されている.カエル胚細胞にC-カドヘリン(カエル胚細胞のAJとデスモソーム両方の構成に関わる)をコートしたビーズを付着させ,細胞間の張力を模した引張力を与えると,細胞間接着部位にプラコグロビンが集積し,繋留されるケラチン繊維も増強されることが示された36).しかし,張力によるプラコグロビンの集積機構など,メカノセンシングの分子機構は明らかとなっていない.一方で,上皮組織自体が発生する張力はAJのアクトミオシン系依存的であり,デスモソームは寄与しないことも示されている37).このことからメカニカルストレスは未分化の上皮細胞におけるデスモソームの構築過程にのみ影響を及ぼすという可能性が考えられるが,デスモソームの結合の安定性は多様な細胞内シグナルで変化するため38),分化した上皮細胞でもメカニカルストレスがデスモソームの安定性を制御している可能性も否定できない.
3)接着斑とメカノセンシング
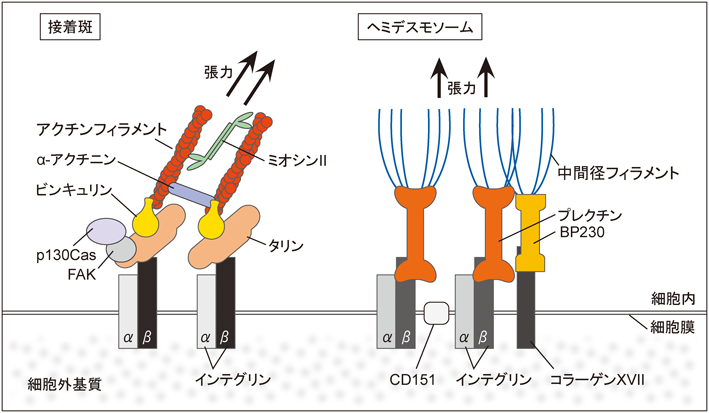
接着斑は結合組織にも上皮細胞にもみられる細胞–基質間接着複合体である.その主要な構成因子であるインテグリンはα鎖とβ鎖のヘテロ二量体からなる膜貫通タンパク質で,β鎖は細胞外で細胞外基質と結合し,細胞内でタリンをはじめとするさまざまな構成タンパク質を介してアクチンストレスファイバーを接着部位に繋留している(図3左).個々の接着斑はおよそ10 nNの力を基質に負荷していることが,動脈平滑筋細胞より単離したストレスファイバーを用いた計測や39),線維芽細胞をシリコーンで形成したマイクロパターン上に接着させた際のシリコーンの歪みの計測によって示されている40).接着斑は単にストレスファイバーによる張力を基質に伝えるだけではなく,その形成や発達は張力による影響を受け,張力と接着斑の面積は比例関係にある40).また,接着斑の増大やストレスファイバーの増加は細胞外基質の硬さ依存的であることが示されており,接着斑が基質の硬さを感知するメカノセンサー機能を持つと考えられている41, 42).
接着斑におけるメカノセンサー分子の探索は盛んに行われており,他の接着複合体と比較すると研究が進んでいる.このうちメカノセンサー機能を持つと最初に報告されたのはp130Casである.p130CasはSrcファミリーチロシンキナーゼの基質であり,FAKと結合することで接着斑に局在する.リン酸化に伴ってアダプタータンパク質Crk, RacのGDP/GTP交換因子DOCK180と三者複合体を形成し,DOCK180によるRacの活性化を亢進させる43).精製したp130Casタンパク質のN末端とC末端をラバー上に固定し引き伸ばす実験によって,p130Casは分子の引き伸ばしによって基質ドメインが露出し,Srcによるリン酸化が亢進することが示された44).しかしp130CasはN末端でFAKと接合するが,C末端と結合するタンパク質は同定されておらず,細胞内でメカノセンサーとして機能しているかについては今後の検証が待たれる.
インテグリンは前述したように細胞と細胞外基質を直接つなぐ接着斑の主要構成因子である.インテグリンの接着活性は細胞内シグナルによる制御を受けており,インテグリンβ鎖に接着斑タンパク質であるタリンが結合するとインテグリンが構造変化して活性化し,細胞外基質との接着が亢進する45, 46).タリンによる細胞内からの制御機構に加えて,細胞外からの引張力やストレスファイバーによる張力によっても,細胞外基質に対するインテグリンの親和性が高まることが知られている.精製タンパク質を用いて結合力を評価すると,インテグリンα5β1は10~30 pNの張力が負荷されると,細胞外基質であるフィブロネクチンとの結合寿命が延びることが示された47).張力の負荷で受容体とリガンドの親和性が高まる現象はキャッチボンドと呼ばれており,真核生物ではインテグリンとフィブロネクチンの結合の他にもP-セレクチンとそのリガンドPSGL-1の結合が知られている48).インテグリンは自身のもつキャッチボンドの性質によって,メカノセンサーとして機能している可能性がある.
タリンは接着斑においてインテグリンとストレスファイバーをつなぐ分子である.タリンは分子内にビンキュリン結合部位を多数保有する.ビンキュリンはアクチン繊維やα-アクチニンと結合するので,タリンとビンキュリンの結合は接着斑やそこに繋留されるストレスファイバーを増強する.タリンのビンキュリン結合部位は通常は分子内相互作用により隠されているが,N末端とC末端を引き伸ばすとビンキュリンの結合が促進することが精製タンパク質を用いて示されている49).また,細胞内でタリンがインテグリンとアクチン繊維をつなぐように配向することや50),タリンの構造変化はミオシンII依存的に生じることも示されており51),タリンは張力依存的な接着斑の増強においてメカノセンサーとして機能している可能性が高いと考えられる.
4)ヘミデスモソームとメカノセンシング
ヘミデスモソームは上皮特異的な細胞–基質間接着複合体であり,インテグリンα6β4が膜貫通タンパク質として細胞外基質と結合している.重層上皮では同じく膜貫通タンパク質であるコラーゲンXVIIも加わりヘミデスモソームが強化される.細胞内ではインテグリンβ4はプレクチンと,コラーゲンXVIIはBP230と結合し,これらがケラチン繊維を接着部位に繋留している(図3右).ヘミデスモソームは上皮細胞を基底膜に連結する役割を持ち,上皮組織のバリア機能に重要であることが古くから知られているが,さらにメカノセンシング機能についての報告も増えている.線虫の発生過程において,筋組織が発生する張力依存的に筋組織と基底膜をつなぐヘミデスモソームの再構築と成熟がみられる52).このときヘミデスモソームに発生した張力によってアダプタータンパク質であるGIT-1がヘミデスモソームにリクルートされ,それによってPIX1-Rac-PAK-1を介した中間径フィラメントのリン酸化が促進される53).また,乳腺上皮細胞は基質が硬い環境では形質転換し間葉系細胞の性質を獲得することが知られているが,このような基質の硬さ依存的な形質転換にはインテグリンα6β4のクラスター形成の抑制,ヘミデスモソーム形成の抑制と,それに伴うPI3K, Racシグナル経路の活性化が関与していることが示された2).上記の例からもメカニカルストレス応答におけるヘミデスモソームの関与は明らかだが,ヘミデスモソームにおけるメカノセンサーの分子実体はほとんど不明である.プレクチンはインテグリンと中間径フィラメントをつなぐ役割を持ち,分子内に力負荷で構造変化することが知られるスペクトリンリピートを持つことから,メカノセンサーとして機能している可能性が考えられる.プレクチン分子のスペクトリンリピートの一部はSH3ドメインと分子内結合していることから,張力負荷によってプレクチンの構造が変化すると,SH3ドメインが露出し,プレクチンがSH3ドメインを介して他のタンパク質と結合できるようになるという可能性が指摘されている54, 55).
4. メカノセンシングにおけるRhoシグナルの関与
低分子量Gタンパク質Rhoファミリー(Rho, Rac, Cdc42など)はアクチン骨格の再構築において重要な役割を担っている56).機械的シグナルによってRhoAの活性化やストレスファイバー形成の促進が認められることから,メカノセンシングにおけるRhoシグナルの関与が注目されている7, 57).Rhoファミリーの活性はGDPとGTPの結合によって制御されており,GDP-GTP交換因子(Rho-GEF)の働きにより不活性型のGDP結合型から活性型のGTP結合型に変換され,活性化される.本節では機械的刺激による細胞骨格の再構築におけるRho-GEFの役割について,我々の知見を中心に紹介する.
1)メカノセンシングに関与するRho-GEFの同定
Rho-GEFは,Dblホモロジードメインを持つDblファミリーと,Dockホモロジー領域を持つDockファミリーに分類される.ヒトでは70種類のDblファミリーと11種類のDockファミリーが存在する.20種類のRhoファミリーメンバーに対して多様なRho-GEFが存在することによって,細胞骨格の多様な制御が可能になっていると考えられる58).DblファミリーのGEF-H1とLargは,引張刺激の下流で,それぞれ異なるシグナル経路で活性化され,引張刺激依存的なRhoAの活性化に関与することが示されている57).
機械的刺激により活性化されるRho-GEFは他にも多く存在すると考えられるが,それを直接証明した報告はきわめて少ない.我々はメカノセンシングに関与するRho-GEFを同定するため,DblファミリーのGEFをそれぞれ発現抑制するshRNAプラスミドライブラリーを作製し,メカノセンシングに関わるRho-GEFのスクリーニングを行った.血管内皮細胞は繰り返し伸展刺激を与えると,伸展方向とは垂直な方向に細胞の長軸とストレスファイバーを配向させる性質がある.ヒト臍帯静脈内皮細胞に各Rho-GEFのshRNAを導入し,繰り返し伸展刺激を与え,刺激後の細胞やストレスファイバーの再配向が影響を受けるかを観察した結果,GEF-H1, Larg, Soloを含む11種類のRho-GEFがこの応答に関与することを見いだした59).これら11種類のRho-GEFはそれぞれ多様なドメイン構造を持ち,標的となるRhoファミリーも異なっており,繰り返し伸展刺激によるストレスファイバー再配向の制御には,複数の多様なRho-GEFが関与していることが示唆された.
2)Rho-GEF Soloと中間径フィラメントによるメカノセンシング機構
細胞に力学的負荷が加わった際,応力は細胞間や細胞–基質間接着部位に最も強く発生する.上記のスクリーニングで見いだした11種類のRho-GEFを血管内皮細胞に発現させ,局在を観察したところ,Solo(ARHGEF40)は細胞間や細胞–基質間に強く局在し,その場所にアクチンが集積することが明らかとなった59).SoloはRhoA/RhoCに対するGEFであり60),ゼブラフィッシュではSoloの相同分子であるQuattroが胚発生の原腸陥入時の収れん伸長運動に関わることが報告されているが61),その機能はほとんど不明であった.
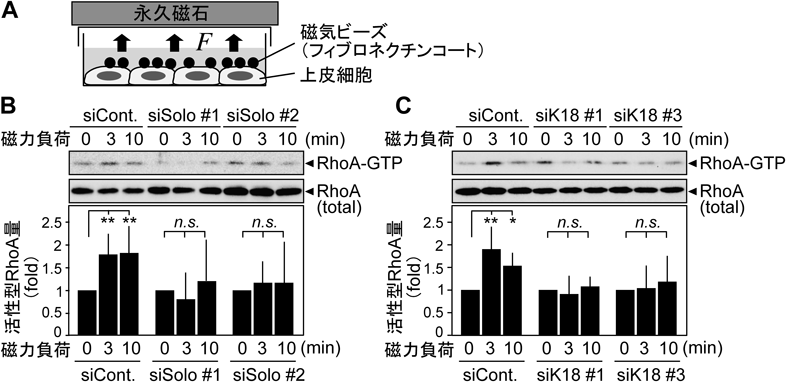
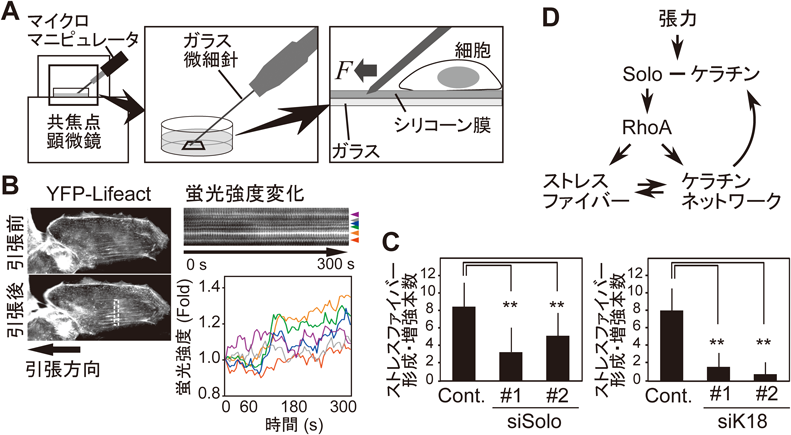
我々は,Soloの結合タンパク質として,上皮細胞における主要な中間径フィラメントであるケラチン8/18を同定した30).さらに,Soloは少なくとも3か所のケラチン結合領域を持つことを示した.次に,引張刺激によるRhoAの活性化に対するSoloやケラチンの関与を検証した.シート状に培養した上皮細胞にフィブロネクチンをコートした磁気ビーズを接着させ,磁石によって細胞に張力を負荷するとRhoAが活性化される(図4).Soloやケラチンを発現抑制すると,RhoAの活性化が抑制され,両者がともに張力負荷依存的なRhoAの活性化に関与することがわかった.また,細胞は張力を負荷されると,その方向にストレスファイバーを形成,増強させる.このような応答をリアルタイムで可視化解析するため,ガラスボトムディッシュに薄いシリコーン膜を張り,フィブロネクチンでコーティングした上に細胞を接着させ,シリコーン膜を針で引っぱって変形させることで細胞に引張刺激を負荷する系を構築した(図5).蛍光タンパク質でアクチンを可視化した上皮細胞に引張刺激を負荷したところ,Soloやケラチンの発現抑制によって,引張刺激で新生・増強されたストレスファイバーの数が抑制されることがわかった.以上の結果から,Soloとケラチン繊維は,引張刺激依存的なRhoAの活性化とストレスファイバーの強化に関与することが明らかとなった30)(図5D).力刺激によるSoloの活性化機構は不明であるが,Soloが複数か所でケラチン繊維と結合することから,ケラチン繊維の構造変化によってSoloが立体構造変化し活性化する可能性が考えられる.
力学的環境に対する細胞や組織の応答機構の解明を目指す「メカノバイオロジー」と呼ばれる融合研究領域が,バイオエンジニアリングと細胞生物学的手法の発展に伴い急速に進展している.細胞–基質間に発生する張力の計測は蛍光ビーズ法(蛍光ビーズを包埋したゲル上に細胞を培養し,ビーズの動きからゲルの歪みを計測する)が最も利用されており,これまでに接着斑およびストレスファイバーのサイズや局在が細胞の張力と相関することが示されている62).さらに近年,細胞接着部位や細胞骨格に発生する力をモニターする張力センサープローブの報告が相次いでいる.張力で構造変化すると想定されているタンパク質の内部に張力で構造変化する弾性リンカーを組み込み,タンパク質の構造変化を両末端に付加した蛍光タンパク質のFRET (fluorescence resonance energy transfer)で検出する手法である.これまでに,ストレスファイバーなどのアクトミオシン系の張力プローブとしてアクチン結合タンパク質のα-アクチニン,フィラミン,スペクトリン,また接着部位の張力プローブとしてカドヘリン,PECAM-1,ビンキュリンを用いたプローブの報告がある63).これらのプローブは多細胞系でも適用例があり,また他のタンパク質にも応用が可能である.こうした力の測定と細胞骨格変化を生細胞で同時計測可能な技術がさらに進展することによって,メカニカルストレスによる細胞骨格制御の分子機構のさらなる解明が期待される.
我々の体を構成するすべての細胞がメカニカルストレスを受けており,適切な力覚応答によって恒常性を維持している.その破綻は多くの病気の原因となっていると予想される.メカノセンシング機構の解明が進むことによって,これまで化学的シグナルだけでは説明できなかった問題について多くの新たな知見が生まれ,さまざまな疾患の病因解明,診断,治療や健康の維持に役立つことが期待される.
引用文献References
1) Hahn, C. & Schwartz, M.A. (2009) Nat. Rev. Mol. Cell Biol., 10, 53–62.
2) Chaudhuri, O., Koshy, S.T., Branco da Cunha, C., Shin, J.W., Verbeke, C.S., Allison, K.H., & Mooney, D.J. (2014) Nat. Mater., 13, 970–978.
3) Iscla, I. & Blount, P. (2012) Biophys. J., 103, 169–174.
4) Ranade, S.S., Syeda, R., & Patapoutian, A. (2015) Neuron, 87, 1162–1179.
5) Higashida, C., Kiuchi, T., Akiba, Y., Mizuno, H., Maruoka, M., Narumiya, S., Mizuno, K., & Watanabe, N. (2013) Nat. Cell Biol., 15, 395–405.
6) Olson, E.N. & Nordheim, A. (2010) Nat. Rev. Mol. Cell Biol., 11, 353–365.
7) Miralles, F., Posern, G., Zaromytidou, A.I., & Treisman, R. (2003) Cell, 113, 329–342.
8) Halder, G., Dupont, S., & Piccolo, S. (2012) Nat. Rev. Mol. Cell Biol., 13, 591–600.
9) Pellegrin, S. & Mellor, H. (2007) J. Cell Sci., 120, 3491–3499.
10) Kreis, T.E. & Birchmeier, W. (1980) Cell, 22, 555–561.
11) Burridge, K. (1981) Nature, 294, 691–692.
12) Hayakawa, K., Tatsumi, H., & Sokabe, M. (2011) J. Cell Biol., 195, 721–727.
13) McGough, A., Pope, B., Chiu, W., & Weeds, A. (1997) J. Cell Biol., 138, 771–781.
14) Uyeda, T.Q., Iwadate, Y., Umeki, N., Nagasaki, A., & Yumura, S. (2011) PLoS ONE, 6, e26200.
15) Tsaturyan, A.K., Koubassova, N., Ferenczi, M.A., Narayanan, T., Roessle, M., & Bershitsky, S.Y. (2005) Biophys. J., 88, 1902–1910.
16) Galkin, V.E., Orlova, A., & Egelman, E.H. (2012) Curr. Biol., 22, R96–R101.
17) Van der Vaart, B., Akhmanova, A., & Straube, A. (2009) Biochem. Soc. Trans., 37, 1007–1013.
18) Janke, C. & Bulinski, J.C. (2011) Nat. Rev. Mol. Cell Biol., 12, 773–786.
19) Desai, A. & Mitchison, T.J. (1997) Annu. Rev. Cell Dev. Biol., 13, 83–117.
20) Kaverina, I., Krylyshkina, O., Beningo, K., Anderson, K., Wang, Y.L., & Small, J.V. (2002) J. Cell Sci., 115, 2283–2291.
21) O’Hagan, R., Chalfie, M., & Goodman, M.B. (2005) Nat. Neurosci., 8, 43–50.
22) Orr, A.W., Helmke, B.P., Blackman, B.R., & Schwartz, M.A. (2006) Dev. Cell, 10, 11–20.
23) Kölsch, A., Windoffer, R., Würflinger, T., Aach, T., & Leube, R.E. (2010) J. Cell Sci., 123, 2266–2272.
24) Snider, N.T. & Omary, M.B. (2014) Nat. Rev. Mol. Cell Biol., 15, 163–177.
25) Galou, M., Gao, J., Humbert, J., Mericskay, M., Li, Z., Paulin, D., & Vicart, P. (1997) Biol. Cell, 89, 85–97.
26) Coulombe, P.A. & Wong, P. (2004) Nat. Cell Biol., 6, 699–706.
27) Kreplak, L., Bär, H., Leterrier, J.F., Herrmann, H., & Aebi, U. (2005) J. Mol. Biol., 354, 569–577.
28) Wang, J. & Pelling, A.(2012) ISRN Cell Biol., 2012, Article ID 513546.
29) Johnson, C.P., Tang, H.Y., Carag, C., Speicher, D.W., & Discher, D.E. (2007) Science, 317, 663–666.
30) Fujiwara, S., Ohashi, K., Mashiko, T., Kondo, H., & Mizuno, K. (2016) Mol. Biol. Cell, 27, 954–966.
31) Maruthamuthu, V., Sabass, B., Schwarz, U.S., & Gardel, M.L. (2011) Proc. Natl. Acad. Sci. USA, 108, 4708–4713.
32) DuFort, C.C., Paszek, M.J., & Weaver, V.M. (2011) Nat. Rev. Mol. Cell Biol., 12, 308–319.
33) Chu, Y.S., Thomas, W.A., Eder, O., Pincet, F., Perez, E., Thiery, J.P., & Dufour, S. (2004) J. Cell Biol., 167, 1183–1194.
34) le Duc, Q., Shi, Q., Blonk, I., Sonnenberg, A., Wang, N., Leckband, D., & de Rooij, J. (2010) J. Cell Biol., 189, 1107–1115.
35) Yonemura, S., Wada, Y., Watanabe, T., Nagafuchi, A., & Shibata, M. (2010) Nat. Cell Biol., 12, 533–542.
36) Weber, G.F., Bjerke, M.A., & DeSimone, D.W. (2012) Dev. Cell, 22, 104–115.
37) Harris, A.R., Daeden, A., & Charras, G.T. (2014) J. Cell Sci., 127, 2507–2517.
38) Garrod, D. & Chidgey, M. (2008) Biochim. Biophys. Acta, 1778, 572–587.
39) Deguchi, S., Ohashi, T., & Sato, M. (2006) J. Biomech., 39, 2603–2610.
40) Balaban, N.Q., Schwarz, U.S., Riveline, D., Goichberg, P., Tzur, G., Sabanay, I., Mahalu, D., Safran, S., Bershadsky, A., Addadi, L., & Geiger, B. (2001) Nat. Cell Biol., 3, 466–472.
41) Geiger, B., Spatz, J.P., & Bershadsky, A.D. (2009) Nat. Rev. Mol. Cell Biol., 10, 21–33.
42) Prager-Khoutorsky, M., Lichtenstein, A., Krishnan, R., Rajendran, K., Mayo, A., Kam, Z., Geiger, B., & Bershadsky, A.D. (2011) Nat. Cell Biol., 13, 1457–1465.
43) Gu, J., Sumida, Y., Sanzen, N., & Sekiguchi, K. (2001) J. Biol. Chem., 276, 27090–27097.
44) Sawada, Y., Tamada, M., Dubin-Thaler, B.J., Cherniavskaya, O., Sakai, R., Tanaka, S., & Sheetz, M.P. (2006) Cell, 127, 1015–1026.
45) Hynes, R.O. (2002) Cell, 110, 673–687.
46) Tadokoro, S., Shattil, S.J., Eto, K., Tai, V., Liddington, R.C., de Pereda, J.M., Ginsberg, M.H., & Calderwood, D.A. (2003) Science, 302, 103–106.
47) Kong, F., García, A.J., Mould, A.P., Humphries, M.J., & Zhu, C. (2009) J. Cell Biol., 185, 1275–1284.
48) Hertig, S. & Vogel, V. (2012) Curr. Biol., 22, R823–R825.
49) del Rio, A., Perez-Jimenez, R., Liu, R., Roca-Cusachs, P., Fernandez, J.M., & Sheetz, M.P. (2009) Science, 323, 638–641.
50) Kanchanawong, P., Shtengel, G., Pasapera, A.M., Ramko, E.B., Davidson, M.W., Hess, H.F., & Waterman, C.M. (2010) Nature, 468, 580–584.
51) Margadant, F., Chew, L.L., Hu, X., Yu, H., Bate, N., Zhang, X., & Sheetz, M. (2011) PLoS Biol., 9, e1001223.
52) Bosher, J.M., Hahn, B.S., Legouis, R., Sookhareea, S., Weimer, R.M., Gansmuller, A., Chisholm, A.D., Rose, A.M., Bessereau, J.L., & Labouesse, M. (2003) J. Cell Biol., 161, 757–768.
53) Zhang, H., Landmann, F., Zahreddine, H., Rodriguez, D., Koch, M., & Labouesse, M. (2011) Nature, 471, 99–103.
54) Ortega, E., Buey, R.M., Sonnenberg, A., & de Pereda, J.M. (2011) J. Biol. Chem., 286, 12429–12438.
55) Osmani, N. & Labouesse, M. (2015) Curr. Opin. Cell Biol., 32, 30–38.
56) Jaffe, A.B. & Hall, A. (2005) Annu. Rev. Cell Dev. Biol., 21, 247–269.
57) Guilluy, C., Swaminathan, V., Garcia-Mata, R., O’Brien, E.T., Superfine, R., & Burridge, K. (2011) Nat. Cell Biol., 13, 722–727.
58) Cook, D.R., Rossman, K.L., & Der, C.J. (2014) Oncogene, 33, 4021–4035.
59) Abiko, H., Fujiwara, S., Ohashi, K., Hiatari, R., Mashiko, T., Sakamoto, N., Sato, M., & Mizuno, K. (2015) J. Cell Sci., 128, 1683–1695.
60) Curtis, C., Hemmeryckx, B., Haataja, L., Senadheera, D., Groffen, J., & Heisterkamp, N. (2004) Mol. Cancer, 3, 10.
61) Daggett, D.F., Boyd, C.A., Gautier, P., Bryson-Richardson, R.J., Thisse, C., Thisse, B., Amacher, S.L., & Currie, P.D. (2004) Curr. Biol., 14, 1632–1638.
62) Schwarz, U.S. & Gardel, M.L. (2012) J. Cell Sci., 125, 3051–3060.
63) Cost, A.L., Ringer, P., Chrostek-Grashoff, A., & Grashoff, C. (2015) Cell. Mol. Bioeng., 8, 96–105.
著者紹介Author Profile
 藤原 佐知子(ふじわら さちこ)
藤原 佐知子(ふじわら さちこ)日本学術振興会特別研究員PD(東北大学大学院生命科学研究科,16年4月より大阪大学大学院基礎工学研究科).博士(生命科学).
略歴2005年東北大学理学部生物学科卒.07年同大学大学院生命科学研究科修士課程修了.07~12年キッセイ薬品工業(株)研究員.15年東北大学大学院生命科学研究科博士課程修了.同年より現職.
研究テーマと抱負機械的刺激(メカニカルストレス)による細胞骨格制御の分子機構について研究を行っています.細胞や組織がどのように力を感じ応答するのか,その実態の理解を目指しています.
趣味バードウォッチング.
 大橋 一正(おおはし かずまさ)
大橋 一正(おおはし かずまさ)東北大学大学院生命科学研究科准教授.博士(理学).
略歴1991年九州大学理学部生物学科卒.96年同大学大学院理学研究科生物学専攻博士課程修了.同年日本学術振興会特別研究員PD. 99年東北大学理学研究科助手.2001年より現職.
研究テーマと抱負細胞が機械的な力を関知してアクチン骨格を再構築するメカニカルストレス応答の分子メカニズムを研究しています.細胞集団の秩序化におけるメカニカルストレス応答の役割を明らかにすることを目指しています.
趣味子供と遊ぶ,熱帯魚飼育.
 水野 健作(みずの けんさく)
水野 健作(みずの けんさく)東北大学大学院生命科学研究科教授.理学博士.
略歴1975年大阪大学理学部卒.79年大阪大学大学院理学研究科博士課程中退.同年宮崎医科大学助手.89年カリフォルニア大学サンディエゴ校博士研究員.90年九州大学理学部助教授.99年東北大学大学院理学研究科教授.2001年より現職.
研究テーマと抱負細胞骨格,力覚応答,繊毛形成を制御するシグナル伝達機構.自分のおもしろいと思った研究を,できるだけ楽しくやっていきたい.
ウェブサイトhttp://www.biology.tohoku.ac.jp/lab-www/mizuno_lab/
趣味テニス,山登り,読書.