1) Grishanin, R.N., Kowalchyk, J.A., Klenchin, V.A., Ann, K., Earles, C.A., Chapman, E.R., Gerona, R.R., & Martin, T.F. (2004) Neuron, 43, 551–562.
2) Speidel, D., Bruederle, C.E., Enk, C., Voets, T., Varoqueaux, F., Reim, K., Becherer, U., Fornai, F., Ruggieri, S., Holighaus, Y., Weihe, E., Bruns, D., Brose, N., & Rettig, J. (2005) Neuron, 46, 75–88.
3) Jockusch, W.J., Speidel, D., Sigler, A., Sørensen, J.B., Varoqueaux, F., Rhee, J.S., & Brose, N. (2007) Cell, 131, 796–808.
4) Sadakata, T., Shinoda, Y., Sekine, Y., Saruta, C., Itakura, M., Takahashi, M., & Furuichi, T. (2010) J. Biol. Chem., 285, 38710–38719.
5) Sadakata, T., Sekine, Y., Oka, M., Itakura, M., Takahashi, M., & Furuichi, T. (2012) FEBS J., 279, 384–394.
6) Sadakata, T., Mizoguchi, A., Sato, Y., Katoh-Semba, R., Fukuda, M., Mikoshiba, K., & Furuichi, T. (2004) J. Neurosci., 24, 43–52.
7) Sadakata, T., Itakura, M., Kozaki, S., Sekine, Y., Takahashi, M., & Furuichi, T. (2006) J. Comp. Neurol., 495, 735–753.
8) Sadakata, T., Kakegawa, W., Mizoguchi, A., Washida, M., Katoh-Semba, R., Shutoh, F., Okamoto, T., Nakashima, H., Kimura, K., Tanaka, M., Sekine, Y., Itohara, S., Yuzaki, M., Nagao, S., & Furuichi, T. (2007) J. Neurosci., 27, 2472–2482.
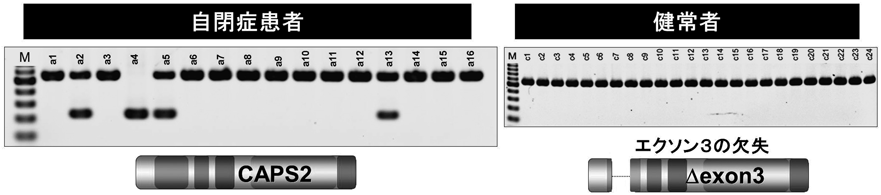
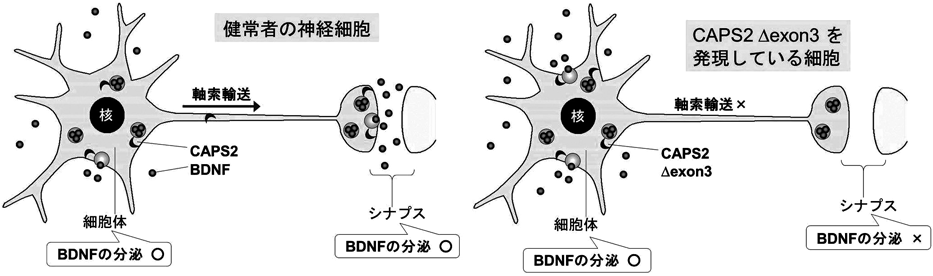
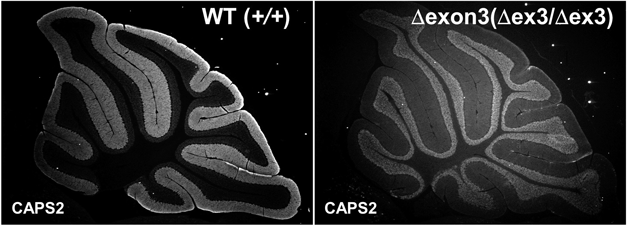
9) Sadakata, T., Washida, M., Iwayama, Y., Shoji, S., Sato, Y., Ohkura, T., Katoh-Semba, R., Nakajima, M., Sekine, Y., Tanaka, M., Nakamura, K., Iwata, Y., Tsuchiya, K.J., Mori, N., Detera-Wadleigh, S.D., Ichikawa, H., Itohara, S., Yoshikawa, T., & Furuichi, T. (2007) J. Clin. Invest., 117, 931–943.
10) Sadakata, T., Shinoda, Y., Oka, M., Sekine, Y., Sato, Y., Saruta, C., Miwa, H., Tanaka, M., Itohara, S., & Furuichi, T. (2012) Proc. Natl. Acad. Sci. USA, 109, 21104–21109.
11) Sadakata, T., Kakegawa, W., Shinoda, Y., Hosono, M., Katoh-Semba, R., Sekine, Y., Sato, Y., Saruta, C., Ishizaki, Y., Yuzaki, M., Kojima, M., & Furuichi, T. (2014) PLoS ONE, 9, e99524.
13) Christian, S.L., Brune, C.W., Sudi, J., Kumar, R.A., Liu, S., Karamohamed, S., Badner, J.A., Matsui, S., Conroy, J., McQuaid, D., Gergel, J., Hatchwell, E., Gilliam, T.C., Gershon, E.S., Nowak, N.J., Dobyns, W.B., & Cook, E.H. Jr. (2008) Biol. Psychiatry, 63, 1111–1117.
14) Sadakata, T., Shinoda, Y., Oka, M., Sekine, Y., & Furuichi, T. (2013) FEBS Lett., 587, 54–59.