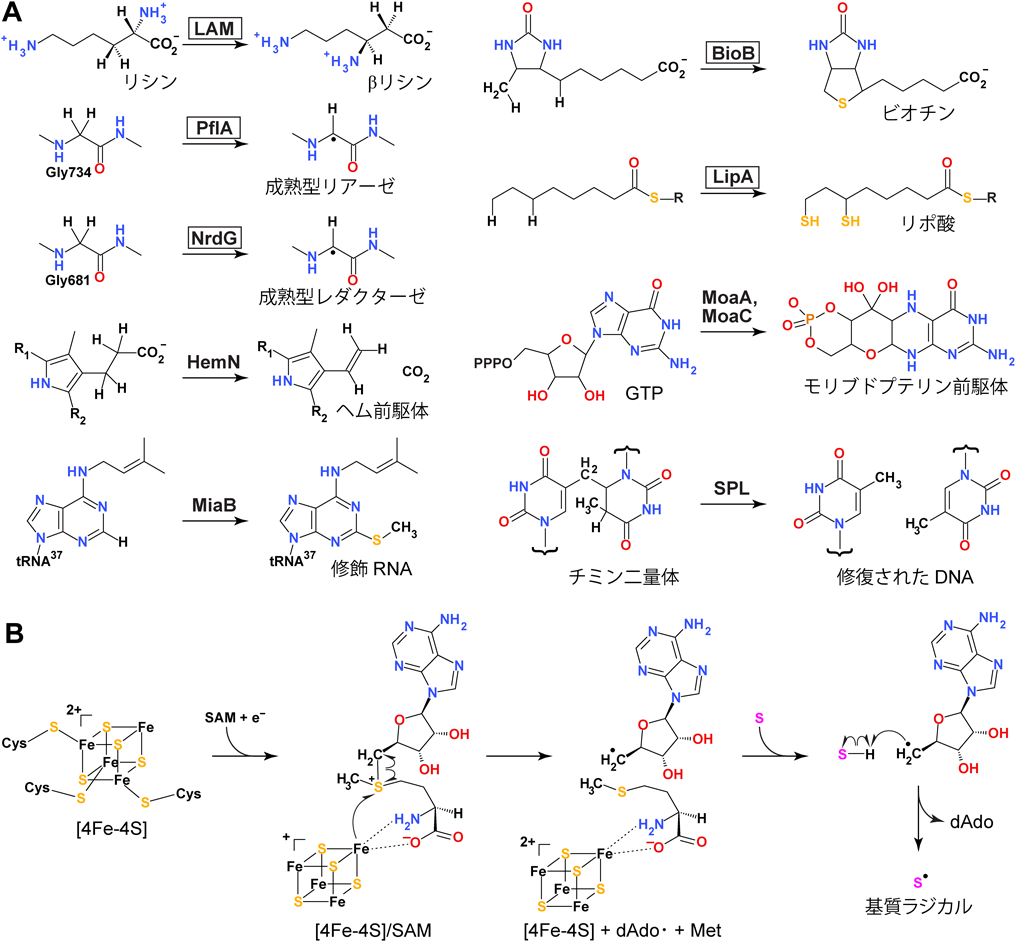
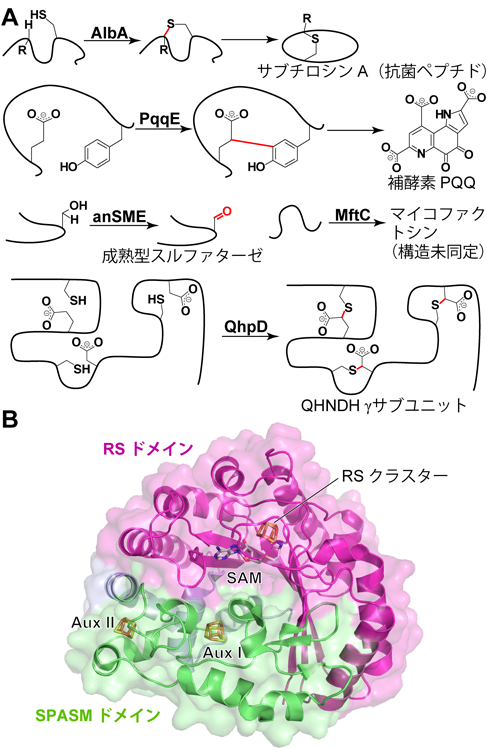
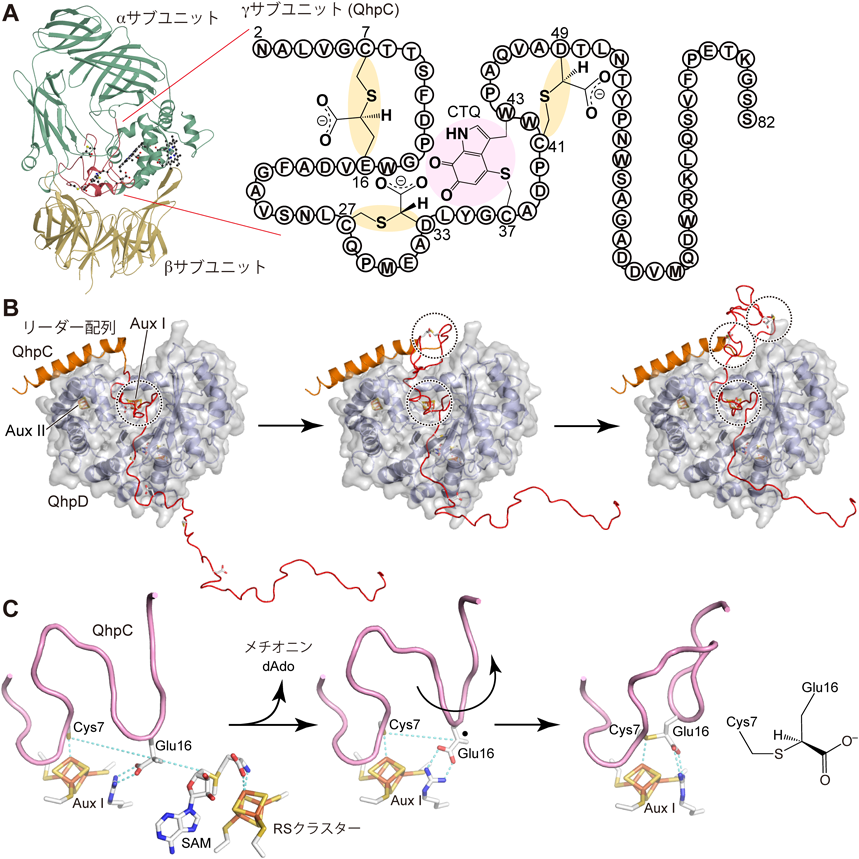
ペプチドを分子内架橋する新規ラジカルSAM酵素Novel radical SAM enzyme forming intrapeptidyl crosslinks
大阪大学産業科学研究所Institute of Scientific and Industrial Research, Osaka University ◇ 〒567–0047 大阪府茨木市美穂ヶ丘8–1 ◇ 8–1 Mihogaoka, Ibaraki, Osaka 567–0047, Japan
発行日:2016年8月25日Published: August 25, 2016