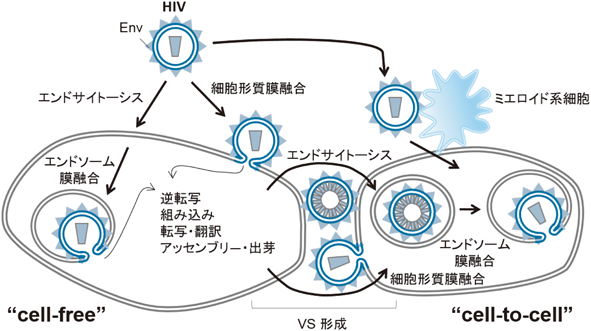
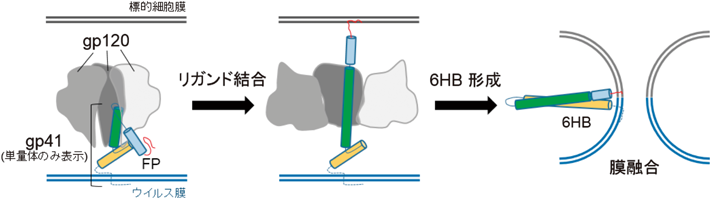
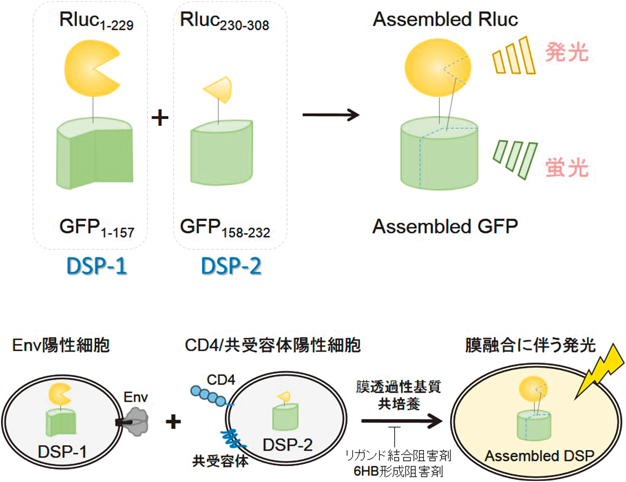
2) Sewald, X., Ladinsky, M.S., Uchil, P.D., Beloor, J., Pi, R., Herrmann, C., Motamedi, N., Murooka, T.T., Brehm, M.A., Greiner, D.L., Shultz, L.D., Mempel, T.R., Bjorkman, P.J., & Priti Kumar, P. (2015) Science, 350, 563–567.
3) Iwami, S., Takeuchi, J.S., Nakaoka, S., Mammano, F., Clavel, F., Inaba, H., Kobayashi, T., Misawa, N., Aihara, K., Koyanagi, Y., & Sato, K. (2015) eLife, 4, e08150.
4) Wei, B.L., Denton, P.W., O’Neill, E.O., Luo, T., Foster, J.L., & Garcia, J.V. (2005) J. Virol., 79, 5705–5712.
5) Miyauchi, K., Kim, Y., Latinovic, O., Morozov, V., & Melikyan, G.B. (2009) Cell, 137, 433–444.
6) de la Vega, M., Marin, M., Kondo, N., Miyauchi, K., Kim, Y., Epand, R.F., Epand, R.M., & Melikyan, G.B. (2011) Retrovirology, 8, 99.
8) Herold, N., Anders-Ößwein, M., Glass, B., Eckhardt, M., Müller, B., & Kräusslich, H.G. (2014) J. Virol., 88, 13956–13970.
9) Vasiliver-Shamis, G., Tuen, M., Wu, T.W., Starr, T., Cameron, T.O., Thomson, R., Kaur, G., Liu, J., Visciano, M.L., Li, H., Kumar, R., Ansari, R., Han, D.P., Cho, M.W., Dustin, M.L., & Hioe, C.E. (2008) J. Virol., 82, 9445–9457.
10) Choudhuri, K., Llodrá, J., Roth, E.W., Tsai, J., Gordo, S., Wucherpfennig, K.W., Kam, L.C., Stokes, D.L., & Dustin, M.L. (2014) Nature, 507, 118–123.
11) Pancera, M., Zhou, T., Druz, A., Georgiev, I.S., Soto, C., Gorman, J., Huang, J., Acharya, P., Chuang, G.Y., Ofek, G., Stewart-Jones, G.B., Stuckey, J., Bailer, R.T., Joyce, M.G., Louder, M.K., Tumba, N., Yang, Y., Zhang, B., Cohen, M.S., Haynes, B.F., Mascola, J.R., Morris, L., Munro, J.B., Blanchard, S.C., Mothes, W., Connors, M., & Kwong, P.D. (2014) Nature, 514, 455–461.
13) Kondo, N., Miyauchi, K., Meng, F., Iwamoto, A., & Matsuda, Z. (2010) J. Biol. Chem., 285, 14681–14688.
14) Kondo, N., Marin, M., Kim, J.H., Desai, T.M., & Melikyan, G.B. (2015) J. Biol. Chem., 290, 6558–6573.
15) Ishikawa, H., Meng, F., Kondo, N., Iwamoto, A., & Matsuda, Z. (2012) Protein Eng. Des. Sel., 25, 813–820.