DNA/RNA編集研究の新たな眺望
1 金沢大学医薬保健学総合研究域医学系
2 島根大学医学部微生物学講座
発行日:2016年10月25日Published: October 25, 2016
© 2016 公益社団法人日本生化学会© 2016 The Japanese Biochemical Society
RNA編集現象は,ゲノム上の遺伝情報がRNAレベルで塩基置換,挿入,欠失などにより,配列が書き換えられる現象である.この現象は1980年代にトリパノソーマのミトコンドリア遺伝子で確認されて以来,植物も含めてさまざまな真核生物で確認されている.哺乳類ではAPOBEC1やADARが行うRNA編集が知られているが,それらのファミリーメンバーにはDNAを標的にするものが含まれる.近年,ゲノム解析やトランスクリプトーム解析の急速な進歩と相まって,DNA/RNA編集研究は新次元の展開を見せている.本特集号では,RNA編集現象,APOBEC, ADARについて,8人の研究者に最新のトピックを紹介していただいた.
DNA/RNA脱アミノ化酵素は,DNAやRNA内にあるシトシンやアデノシンのアミノ基を酸素分子に換える核酸修飾酵素(これをデアミナーゼと呼ぶ)である.哺乳類では,シトシンを触媒するAPOBECファミリーとアデノシンを触媒するADARファミリーより構成され,これら二者はデアミナーゼスーパーファミリーを形成している(図1,図2参照).このスーパーファミリーの共通する構造上の特性は,Znフィンガーからなるデアミナーゼモチーフを持つことであるが,ファミリー内にはデアミナーゼ活性が確認されていないものもある.
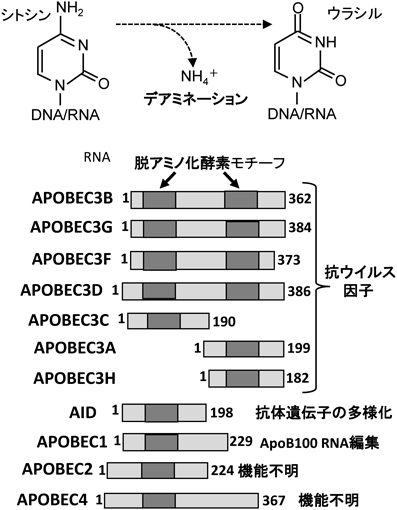
APOBECファミリーは,ヒトでは11種類が知られており,H-X-E-X(23–25)-P-C-X(2–4)-Cのアミノ酸配列からなるシチジンデアミナーゼモチーフを一つないし二つ持つことを共通の生化学的特徴としている(図1).ファミリー名の由来となったAPOBEC1(apolipoprotein B100 RNA editing catalytic subunit-1)は,最初に単離されたファミリーメンバーであり1),哺乳類でRNA編集を行う酵素として最も研究が進んでいるデアミナーゼの一つである.APOBEC1は小腸や肝臓においてapolipoprotein B100のmRNA中のシトシンをウラシルに変換することで,翻訳領域内部に終止コドンを作り,本来ApoB100タンパク質をコードするmRNAを,ApoB48タンパク質をコードするmRNAに変換する.ApoB100とApoB48タンパクは,脂質の輸送においてそれぞれ異なる役割を担うので,APOBEC1は一つのmRNAから2種類の機能の異なるタンパク質を作りだすと言えよう.AIDは,APOBEC1に次いで単離されたデアミナーゼで,抗体遺伝子座で起こるクラススイッチ組換え,somatic hypermutation, gene conversionという遺伝子改編現象を担うことが知られている2).AIDの起こす遺伝子改編現象は,抗体分子の多様化や抗体の親和性成熟につながり,獲得免疫の要と考えられている.APOBEC2は,筋組織に限局した発現パターンを示すAPOBECタンパク質で,筋組織の発生や構築に関わることが示唆されているが,その分子機構はほとんどわかっていない.またAPOBEC4についてもmRNA発現が精巣で確認されているものの機能は不明である1).APOBEC3は,ウイルスDNAを標的とし抗ウイルス効果を発揮する自然免疫効果分子である.その標的ウイルスにHIV-1や発がんウイルスであるパピローマウイルスが含まれることや,がんゲノムの進展との関わりが示唆されていることより3),現在,最も注目されているAPOBECタンパク質といえる.本特集では,5人の研究者より,抗ウイルス活性,構造,進化,病態,発がんなどの見地から,APOBEC3研究の面白さを概説していただいた(村松,高折,佐藤,宮澤,都築の総説を参照).
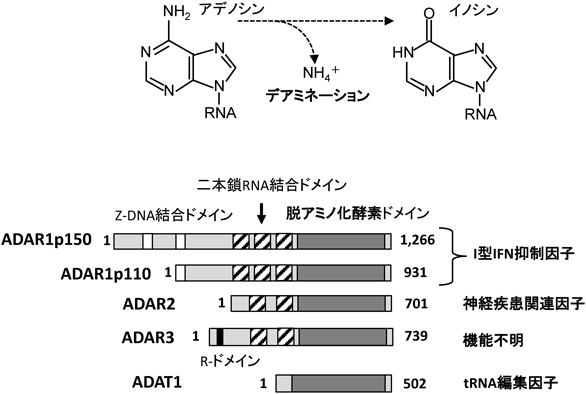
一方,ADARファミリーは,ヒトでは少なくともADAR1~3の3種類が確認されている.ADARファミリーは総じてAPOBECタンパク質よりはるかに大きく,一つ以上の二本鎖RNA結合ドメインと一つのデアミナーゼドメインを持つ4)(図2).
ADAR1は,Z-DNA結合ドメインを二つ持つADAR1p150と,一つ持つADAR1p110があり,両者は同じ遺伝子座から転写開始点の違いにより作られる.ADAR1は全身の組織で発現しているが,ADAR1p150とADAR1p110はそれぞれ異なる細胞内局在をとる.ADAR1p150の発現はインターフェロン-α, β(I型IFN)によって誘導されるが,最近ADAR1はI型IFNシグナルの抑制因子であることが明らかとなった.さらに,ADAR1は,microRNA前駆体と結合し,その産生を制御する.
ADAR2は,神経組織に強く発現し,AMPA型グルタミン酸受容体を構成するサブユニットの一つGluA2にRNA編集を引き起こす.ADAR2は,神経に発現するさまざまなRNAを基質とするが,近年筋萎縮性側索硬化症などの神経変性疾患や,プラダー・ウィーリー症候群を含む精神疾患との関連性が指摘されている.
ADAR3は神経組織に限局して発現し,他のADARと異なりデアミナーゼ活性を持っていない.また,N末端にアルギニン(R)リッチなR-ドメインを持ち一本鎖RNAと結合するが,ADAR3の生理的意義はよくわかってない.
本特集では,2人の研究者より,免疫疾患,がん,microRNA制御,筋萎縮性側索硬化症を中心とした神経疾患などの見地から,ADARについて執筆いただいた(飯笹と山下の総説を参照).
一方,ADARやAPOBECとはまったく異なるRNA編集機構が,植物に存在することが報告されている.植物のRNA編集については,シチジンデアミナーゼ様の配列DYWドメインを含むPRP(pentatricopeptide repeat)タンパク質について,竹中先生に執筆いただいた.
本特集号によりRNA編集現象とDNA/RNA編集研究の面白みが,『生化学』読者に伝わることを切に願っている.
1) Smith, H.C., Bennett, R.P., Kizilyer, A., McDougall, W.M., & Prohaska, K.M. (2012) Semin. Cell Dev. Biol., 23, 258–268.
2) Muramatsu, M., Kinoshita, K., Fagarasan, S., Yamada, S., Shinkai, Y., & Honjo, T. (2000) Cell, 102, 553–563.
3) Harris, R.S. & Dudley, J.P. (2015) Virology, 479–480, 131–145.
4) Nishikura, K. (2016) Nat. Rev. Mol. Cell Biol., 17, 83–96.

This page was created on 2016-08-31T09:10:36.474+09:00
This page was last modified on 2016-10-14T11:58:47.980+09:00
このサイトは(株)国際文献社によって運用されています。