植物オルガネラのRNA編集とDYWドメインDYW domain and RNA editing in plant organelles
ウルム大学分子植物学University Ulm, Molecular Botany ◇ Albert-Einstein-Allee11, D-89069 Ulm, Germany ◇ Albert-Einstein-Allee11, D-89069 Ulm, Germany
発行日:2016年10月25日Published: October 25, 2016
植物ではオルガネラmRNA上のC-to-UおよびU-to-C RNA編集が知られている.これまでに単離された個々のRNA編集サイト特異的な因子は配列特異的なRNA結合タンパク質PPR(pentatricopeptide repeat)タンパク質であり,その約半数がC末端側にDYWドメインというシチジンデアミナーゼ(cytidine deaminase:CDA)モチーフ様配列を保持している.このドメインはHXE(x)nCXXCの保存されたZnイオン結合モチーフを持ち,CDAとして機能するようにみえるがその酵素活性は証明されていない.最近それ以外のRNA編集因子が次々と報告され,これらの因子がDYWドメインを持つPPRタンパク質と複雑に相互作用することが明らかになってきた.これは植物オルガネラでも他の生物のRNA編集と同様,RNA編集タンパク質複合体(エディトソーム)が存在することを示唆している.
© 2015 公益社団法人日本生化学会© 2015 The Japanese Biochemical Society
植物オルガネラのmRNA上で特定のCがUへ変換されるRNA編集はこれまでゼニゴケ以外のすべての陸上植物で観察されている1).RNA編集サイトの数は植物種によって異なり,たとえば蘚類ヒメツリガネゴケではミトコンドリアに11か所,葉緑体では1か所しか存在しないのに対し,ツノゴケ,シダ,セラギネラでは1000か所以上に及ぶ2–4).しかし被子植物ではミトコンドリアで400~600か所,葉緑体では30~50か所と数が減少する.興味深いことにツノゴケやある種のシダではC-to-Uと逆向きのU-to-C RNA編集も存在している1).RNA編集の約80%は翻訳産物のアミノ酸配列を変化させる.また,時には葉緑体ndhD遺伝子のようにACGをAUGに変え開始コドンを作製する.U-to-C RNA編集を持つ種ではDNAにコードされる終止コドンがRNA編集により除去される例も頻繁にみられる2, 3).これらの例はRNA編集が植物オルガネラのタンパク質機能の発現に重要な役割を果たしていることを示している.実際RNA編集変異体の中にはミトコンドリアの機能低下により著しく成長が阻害されたり,葉緑体の発達が阻害されたりといった強い表現型を見せるものが数多く報告されている5, 6).
植物RNA編集機構研究の初期には,オルガネラ抽出タンパク質を用いて試験管内でRNA編集反応を再現するin vitro系が反応の生化学的特徴や反応に必要なシス配列の同定に大いに寄与した.in vitro RNA編集反応後に32Pで標識されたmRNA上のC塩基の一部が32P標識Uに変換されることが示され7),この反応がCのピリミジン基のデアミナーゼ反応であることが明らかになった.さらに配列の一部を欠失,変異させたmRNAをin vitro系で解析することで,RNA編集に必要なシス配列が各サイトの−5から−25塩基の間に含まれることが明らかになった8, 9).
2005年の最初のRNA編集因子CRR4の報告以降10),数多くの植物オルガネラRNA編集因子がシロイヌナズナやヒメツリガネゴケ,トウモロコシ,イネといったモデル植物を用いた遺伝学的手法により単離されてきた11).これらのRNA編集因子の中にはシチジンデアミナーゼ(cytidine deaminase:CDA)様の配列であるDYWドメインを含むPPRタンパク質(pentatricopeptide repeat:PPR)が多数単離されている.しかし,いまだ同定された因子を再構築しRNA編集をin vitroで再現するには至っておらず,その活性酵素は未知のままである.本稿では,これまでに単離されたRNA編集因子,特にCDA様の配列を持つDYWドメインの構造および機能に関する知見を紹介しつつ,エディトソーム(editosome;RNA編集タンパク質複合体)の全体像について考察したい.
葉緑体のndh1サイトのRNA編集に必要な因子として単離されたCRR410),および最初のミトコンドリアRNA編集因子MEF112)はいずれもPPRタンパク質であった.このタンパク質は約35アミノ酸からなる二つのαヘリックス構造を持つPPRモチーフを繰り返し保持している.PPRタンパク質遺伝子は酵母,動物を含む多くの真核生物の核ゲノムにコードされているが特に陸上植物でその数が著しく多い13).たとえば,藻類クラミドモナスでは19個なのに対し,ヒメツリガネゴケでは109個,シロイヌナズナでは496個14)のPPR遺伝子がコードされている.PPRタンパク質はその構造からP, PLSの2クラスに分けられる.Pクラスは35アミノ酸のPPRモチーフが並んだ構造でスプライシング,RNAプロセッシング,翻訳などさまざまなRNA機能調節に関わっている.一方PLSクラスPPRタンパク質の構造は,P(35アミノ酸),L(long, 36~40アミノ酸),S(short, 31アミノ酸)の三つのPPRがユニットを構成し,それが反復した構造を持つ(図1A).このPLSクラスPPRはC末端側のドメインの違いにより,さらに三つのサブクラスに分類される13, 14).C末端側にE(extension)ドメインのみを持つものはEサブクラス,Eに加えてそのC末端にDYWドメイン(C末端の保存された3アミノ酸,Asp, Tyr, Trpにより命名された)が付加されたものはDYWサブクラスに分類される(図1A).これまでに単離された各サイトに特異的なRNA編集因子はすべてEまたはDYWサブクラスに属する.シロイヌナズナにはE, DYWサブクラスのPPRが約100個ずつコードされており,これは400~600のRNA編集サイトを認識するのに十分な数といえる.PPRタンパク質は一般的に一本鎖RNAと特異的に結合するが15, 16),RNA編集因子についても,そのPPRドメインがそれぞれのサイトの5′側の配列(シス配列)と特異的に結合していることが実験的に明らかになってきた17, 18).
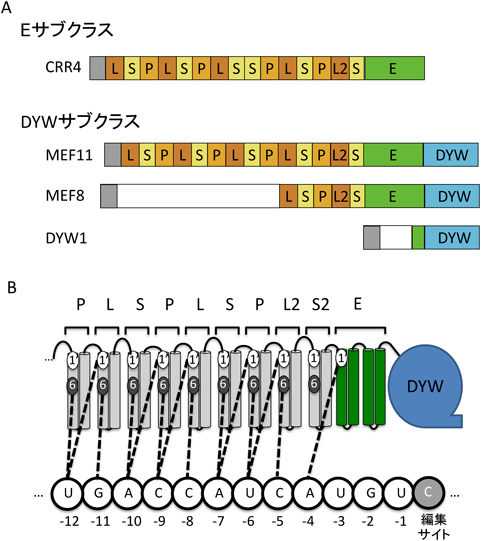
(A)代表的なEサブクラス,DYWサブクラスPPR型のRNA編集因子.ほとんどのRNA編集因子は10個ほどのPPRモチーフを持っている.DYW1, MEF8のPPRドメインは短く,EサブクラスPPRにDYWドメインを供給する因子と思われる(図3を参照).(B)PPRコードでは各PおよびSタイプPPRモチーフの6番目と次のモチーフの1番目(1′)の位置のアミノ酸の組合わせによって認識する塩基が決まる19–21).たとえば,6, 1′がTNの場合Aを認識する.L, L2ドメインでは6番目の位置のアミノ酸のみ,S2ドメイン(最もC末端側のSモチーフ)では1′の位置のアミノ酸のみがそれぞれ認識に関与している.
PPRタンパク質の特異的RNA認識の分子機構について最近画期的な報告がなされた.BarkanらはRNA編集因子の各PPRのモチーフをRNA編集サイトの4塩基上流からさらに上流へ1塩基ごとに並べた場合,Lを除くP, Sモチーフの2か所のアミノ酸(6番目と次のモチーフの最初のアミノ酸1′,注:PPRモチーフ中のアミノ酸の数え方はいくつか提唱されている)の組合わせが,対応するRNA塩基の種類と相関している(つまり塩基認識に重要である)といういわゆるPPRコードを提唱した(図1B)19).さらにその6, 1′の2か所のアミノ酸を置換することにより組換えPPRタンパク質の配列特異性を変化させることに成功し,その仮説が基本的に正しいことを示した19).一方中村らおよび筆者らはそれぞれ独自にP, Sモチーフだけではなく,Lモチーフも塩基認識へ関与している可能性をin silico解析により示した(図1B,中村らはさらに1か所のアミノ酸を加えた3アミノ酸による塩基認識を提唱している)20, 21).
このPPRコードはPLSクラスPPRがいかにそれぞれの編集サイトを認識するかを明快に説明できる.実際PPRのアミノ酸配列から標的となるRNA編集サイトを予測することで,新奇なRNA編集因子がいくつか単離された22, 23).しかしながら現在のPPRのコードでは説明できないRNA編集因子とその標的RNAの組合わせもある20, 21, 24).またその後に解明されたPPRタンパク質の立体構造は,PPRタンパク質コードが基本的には正しいものの,その他にもRNA分子との相互作用に影響するアミノ酸がある可能性を示唆している25).
PPR型RNA編集因子はC末端にEドメインまたはE−DYWドメインを持つ.Eドメインは以前から2個のPPR様モチーフを持っていることが示唆されており,その構造からPPR同様にRNA結合に関与している可能性が示唆されてきた14).しかしながら葉緑体RNA編集因子CRR22を用いた奥田らの解析結果はEドメインがRNA結合に寄与していないことを示唆している26).我々の最近のデータではEドメインが,後述するMORFタンパク質27)という他のRNA編集因子と相互作用することを示唆しており,このドメインはサイト特異的なRNA編集複合体構成上の要である可能性がある.Eドメインの配列は各RNA編集因子で比較的多様であるため,そのRNA結合能,タンパク質結合能については個別の機能解析が必要である.
DYWドメインは,EドメインのC末端側に位置する.植物ではDYWサブクラスPPRの有無とオルガネラにおけるC-to-U RNA編集が常にリンクしていること28),DYWサブクラスPPRを遺伝子水平移行により獲得したと思われるアメーバ,Naegleria gruberiのミトコンドリアが植物型のC-to-U RNA編集を持つこと29)などはいずれもDYWドメインがRNA編集反応に不可欠であることを示唆している.
RNA編集因子に含まれるDYWドメインはCDAに保存されるZn結合ドメインHXE(x)nPCXXCに類似した配列HSE(x)nCXDCを含んでおり(図2),予測される二次構造も既知のCDAとよく似ている30).ただしDYWではCysに隣接したProが保存されていない.DYWドメインへのZnイオン結合能はICP-MS(inductively coupled plasma-mass spectrometry)により確認された31, 32).またDYWドメイン内のHSEとCXDC部位の欠失,または変異によりそのZnイオン結合能も失われ,RNA編集能も失われた31, 32).DYWドメインのHXEモチーフのN末端側には既知のデアミナーゼにはない約40アミノ酸の挿入が存在し,その中には保存されたアミノ酸がいくつかある30).これらのアミノ酸が標的となるCやその周辺のRNAの認識に関わっているのかもしれない.

シロイヌナズナのミトコンドリアRNA編集因子MEF1, MEF8および葉緑体RNA編集因子OTP85, OTP84, DYW1, ヒメツリガネゴケミトコンドリアのPpPPR_71.黒三角はデアミナーゼに保存されるHXE, CXXCモチーフの位置を示す.予想されるヘリックス(Helix),ストレッチ(Str)の位置はIyerらの論文を参照した30).
DYWドメインのC末端側アミノ酸配列は他のデアミナーゼの構造と特に相似していないにも関わらず,高度に保存されている(図2).特に最後のC末端のアミノ酸は重要で,DYW1ではC末端Wのアラニンへの変異,C末端3アミノ酸の欠失により,いずれもそのRNA編集能を失う31).またいくつかのDYWサブクラスRNA編集因子のC末端にポリペプチドを付加すると活性を失ってしまう33, 34).C末端のアミノ酸が特異的なタンパク質結合に必要である例はいくつか知られているが(小胞体にとどまるのに必要なK/HDELシグナル,ペルオキシソーム局在シグナル,SKLなど35)),C末端DYW配列の高度な保存性は,この部位が他のタンパク質との特異的な結合に必要なシグナル配列である可能性を示している.
組換えタンパク質を用いたRNA結合実験により,DYWドメイン自体がRNA結合能を持つこと,またその結合能は標的となるサイトがRNA編集を受ける前のCを含む配列の方が,編集後のUを含む配列よりも高いことが示された36).また奥田らはCRR28, OTP85のDYWドメインのRNA結合場所がRNA編集サイトとその上流3塩基部分に及ぶことを示した26).これらの結果はいずれもDYWドメインがRNA編集サイトに直接結合し,またCDA反応に関係していることを示唆しているが,その酵素活性はいまだ証明されておらず,未知の補因子が存在する可能性を示している.
前述のとおり約半数のPPR型RNA編集因子はDYWドメインを持たない.これは他のタンパク質のDYWドメインがEサブクラスPPRへ供給されているからと考えられる(図3).その例として一番よく知られているのが,CRR4とDYW1である37).Eドメインのみを持つCRR4とDYW1ドメインしか持たないタンパク質DYW1はいずれも葉緑体のndhD-1サイトのRNA編集に必要である.またCRR4とDYW1を連結させたキメラタンパク質は二つの遺伝子の二重変異体を相補することから,CRR4とDYW1のタンパク質複合体がndhD-1サイトのRNA編集に必要であることを示唆している.DYWサブクラスPPRの中にはDYWドメインを欠失させ,PPRとEドメインのみになってもそのRNA編集能を失わないものがある38, 33, 39).この結果はEサブクラスのRNA編集因子ではDYWドメインが他の因子から相補されているという仮説を支持している.
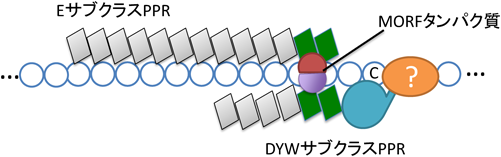
EサブクラスPPRはRNA編集サイトシス配列を認識して結合する.MORFはDYWサブクラスPPRとEサブクラスPPRのタンパク質複合体形成を補助する.未知の因子がDYWドメインのC末端に結合してCDA反応を完遂する.図には示していないが,RRMタンパク質もこの複合体に加わりPPRタンパク質のRNA編集サイトへの結合を補助する.OZ1はDYW近辺に結合し,Znイオンを供給するのかもしれない.
DYWドメインを供給する側の因子についてはDYW1以外にも候補が見つかっている.ミトコンドリアのDYWサブクラスRNA編集因子であるMEF8のPPRモチーフは5個しかなく配列保存性も低いため,塩基配列特異的にRNAに結合しているとは考えにくい.またMEF8変異体の表現型は,RNA編集活性がほぼなくなる他のPPR型RNA編集因子の変異体とは異なり,複数個のRNA編集サイト活性が約50%に低下するものである40).MEF8同様少数のPPRモチーフしか持たないDYWサブクラスPPRはDYW1, MEF8を含めて6個コードされている.これらは他のEサブクラスPPRにDYWを供給する側のPPRタンパク質の第一候補である37, 40).
他の因子によるDYWドメインの相補は常に機能するわけではない.MEF1, OTP85/QED1などはDYWドメインを欠失させるとRNA編集活性を失ってしまう33, 34).OTP84の場合,DYWを欠失させると三つの標的となるサイトのうち二つしかRNA編集できなくなる41).これらの結果はDYWドメインの機能が各々異なっており,本来DYWサブクラスPPRにあったDYWの機能が,他の因子から供給されたDYWドメインでは完全に相補されない場合があることを示している.
筆者らは遺伝学手法によりミトコンドリアで多数のRNA編集サイトに関与するMORF1(multiple organellar RNA editing factor 1),MORF3および,葉緑体で多数のRNA編集サイトに関与するMORF2とMORF9を単離した27).ほぼ同時期に葉緑体のRNA編集因子RARE1と相互作用するRIP1がミトコンドリア,葉緑体双方でRNA編集に関わっている因子として単離されたが,これはMORFと同じファミリーに属するタンパク質である(RIP1=MORF8)42).いくつかのRNA編集サイトが高効率でRNA編集されるためには複数のMORFを必要とする.MORFタンパク質はホモ,ヘテロで相互作用するとともに,PPR型RNA編集因子とも結合することからRNA編集タンパク質複合体の形成に複雑に関与していると思われる27, 43).たとえばEサブクラスPPR型RNA編集タンパク質であるMEF13とMORF3は酵母ツーハイブリッドでほとんど相互作用しないにも関わらず,MORF8を3番目のタンパク質として発現すると著しく強い相互作用を示す44).一方MORF1では同様の効果は観察されなかった.これは特定のMORFタンパク質の組合わせによりPPR型RNA編集因子との相互作用が変化することを示している.筆者らはMORFタンパク質がEサブクラスとDYWサブクラスPPRの選択的タンパク質複合体形成を手助けする役割を持っているのではないかと考えている1)(図3).
RRMドメイン(RNA recognition motif)を持つタンパク質Cp31は最初葉緑体in vitro系でRNA編集を受けるRNAに結合するタンパク質として単離され45),後にそのRNA編集への関与が変異体解析により証明された46).さらに別のRRMタンパク質であるORRM1(Organelle RRM)は葉緑体の,ORRM2, ORRM3, ORRM4はミトコンドリアのRNA編集に関わっていることが示された47–49).これらのRRMタンパク質はPPRやMORFと関与しながらRNAの二次構造を制御することにより効率的なRNA編集に関与していると思われる.
Znイオン結合ドメインを持つOZ1(organelle zinc finger 1)はORRM1と相互作用する因子として単離され,葉緑体RNA編集への関与が明らかにされた50).OZ1はPPRと相互作用するがMORF/RIPタンパク質と直接相互作用していない.OZ1はその構造からDYWドメインへのZnイオン供給に関わっている可能性があるが,詳しい分子機能は明らかになっていない.
最近PクラスPPR, PPMEの変異体でnad1の二つのサイトのRNA編集がほぼ完全に失われることが報告された51).またEドメインを持たないPLSクラスPPRであるPDM1/SEL1はMORFタンパク質と相互作用を示し,効率的なRNA編集に関与している52).PPRタンパク質は,編集サイト近辺に結合しRNA二次構造および他のRNA編集因子のRNA結合に影響を与えると思われる.今後もEドメインを保持しないPPRタンパク質の中からRNA編集に関わる因子が発見される可能性は大いにある.
シロイヌナズナのゲノムには他の生物のシチジンデアミナーゼと高い相同性を持つ普遍型シチジンデアミナーゼ(CDA)遺伝子が九つあるが,そのうち酵素活性を持つのは一つだけである.このCDAがオルガネラのRNA編集へ関与する可能性は否定されている53, 54).
RNA編集効率は組織間で異なっており55),ペチュニアではRNA編集前後の2種類のmRNA両方からRPS12タンパク質が翻訳されていることが報告されている56).このことはRNA編集の有無によって生じる異なるアミノ酸配列を持つタンパク質がそれぞれ異なる機能,活性を持ち,オルガネラ機能を制御している可能性を示唆しているが,その明確な証拠は見つかっていない.
近年さまざまな因子がRNA編集に関わる因子として単離され,これらが複合体を形成していることがわかってきた.しかし,まだその全体像ははっきりしていない.最大の課題はこのタンパク質複合体のCDA活性に必要な未知の因子の同定である.遺伝学的またin vitroRNA編集系を利用した生化学的手法を駆使し,CDA活性を持つRNA編集タンパク質複合体を再現する必要がある.CDA様配列を持つDYWドメインはその未知因子を特定する鍵となるであろう.
1) Takenaka, M., Zehrmann, A., Verbitskiy, D., Härtel, B., & Brennicke, A. (2013) Annu. Rev. Genet., 47, 335–352.
2) Kugita, M., Yamamoto, Y., Fujikawa, T., Matsumoto, T., & Yoshinaga, K. (2003) Nucleic Acids Res., 31, 2417–2423.
3) Wolf, P.G., Rowe, C.A., & Hasebe, M. (2004) Gene, 339, 89–97.
4) Oldenkott, B., Yamaguchi, K., Tsuji-Tsukinoki, S., Knie, N., & Knoop, V. (2014) RNA, 20, 1499–1506.
5) Zhu, Q., Dugardeyn, J., Zhang, C., Takenaka, M., Kühn, K., Craddock, C., Smalle, J., Karampelias, M., Denecke, J., Peters, J., Gerats, T., Brennicke, A., Eastmond, P., Meyer, E.H.E.H., Van Der Straeten, D., Kuhn, K., Craddock, C., Smalle, J., Karampelias, M., Denecke, J., Peters, J., Gerats, T., Brennicke, A., Eastmond, P., Meyer, E.H.E.H., & Van Der, S.D. (2012) Plant J., 71, 836–849.
6) Chateigner-Boutin, A.-L., Ramos-Vega, M., Guevara-García, A., Andrés, C., de la Luz Gutiérrez-Nava, M., Cantero, A., Delannoy, E., Jiménez, L.F., Lurin, C., Small, I., & León, P. (2008) Plant J., 56, 590–602.
7) Yu, W. & Schuster, W. (1995) J. Biol. Chem., 270, 18227–18233.
8) Takenaka, M. & Brennicke, A. (2003) J. Biol. Chem., 278, 47526–47533.
9) Miyamoto, T., Obokata, J., & Sugiura, M. (2002) Mol. Cell. Biol., 22, 6726–6734.
10) Kotera, E., Tasaka, M., & Shikanai, T. (2005) Nature, 433, 326–330.
11) Fujii, S. & Small, I. (2011) New Phytol., 191, 37–47.
12) Zehrmann, A., Verbitskiy, D., van der Merwe, J., Brennicke, A., & Takenaka, M. (2009) Plant Cell, 21, 558–567.
13) Schmitz-Linneweber, C. & Small, I. (2008) Trends Plant Sci., 13, 663–670.
14) Cheng, S., Gutmann, B., Zhong, X., Ye, Y., Fisher, M.F., Bai, F., Castleden, I., Song, Y., Song, B., Huang, J., Liu, X., Xu, X., Lim, B.L., Bond, C.S., Yiu, S.-M., & Small, I. (2016) Plant J., 85, 532–547.
15) Nakamura, T., Schuster, G., Sugiura, M., & Sugita, M. (2004) Biochem. Soc. Trans., 32, 571–574.
16) Beick, S., Schmitz-Linneweber, C., Williams-Carrier, R., Jensen, B., & Barkan, A. (2008) Mol. Cell. Biol., 28, 5337–5347.
17) Okuda, K., Nakamura, T., Sugita, M., Shimizu, T., & Shikanai, T. (2006) J. Biol. Chem., 281, 37661–37667.
18) Hammani, K., des Francs-Small, C.C., Takenaka, M., Tanz, S.K., Okuda, K., Shikanai, T., Brennicke, A., & Small, I. (2011) J. Biol. Chem., 286, 21361–21371.
19) Barkan, A., Rojas, M., Fujii, S., Yap, A., Chong, Y.S., Bond, C.S., & Small, I. (2012) PLoS Genet., 8, e1002910.
20) Yagi, Y., Hayashi, S., Kobayashi, K., Hirayama, T., & Nakamura, T. (2013) PLoS ONE, 8, e57286.
21) Takenaka, M., Zehrmann, A., Brennicke, A., & Graichen, K. (2013) PLoS ONE, 8, e65343.
22) Yagi, Y., Tachikawa, M., Noguchi, H., Satoh, S., Obokata, J., & Nakamura, T. (2013) RNA Biol., 10, 1419–1425.
23) Chateigner-Boutin, A.L., Colas Des Francs-Small, C., Fujii, S., Okuda, K., Tanz, S.K., & Small, I. (2013) Plant J., 74, 935–945.
24) Arenas-M, A., Takenaka, M., Moreno, S., Gómez, I., & Jordana, X. (2013) FEBS Lett., 587, 887–891.
25) Yin, P., Li, Q., Yan, C., Liu, Y., Liu, J., Yu, F., Wang, Z., Long, J., He, J., Wang, H.-W., Wang, J., Zhu, J.-K., Shi, Y., & Yan, N. (2013) Nature, 504, 168–171.
26) Okuda, K., Shoki, H., Arai, M., Shikanai, T., Small, I., & Nakamura, T. (2014) Plant J., 80, 870–882.
27) Takenaka, M., Zehrmann, A., Verbitskiy, D., Kugelmann, M., Härtel, B., & Brennicke, A. (2012) Proc. Natl. Acad. Sci. USA, 109, 5104–5109.
28) Rüdinger, M., Polsakiewicz, M., & Knoop, V. (2008) Mol. Biol. Evol., 25, 1405–1414.
29) Rudinger, M., Fritz-Laylin, L., Polsakiewicz, M., & Knoop, V. (2011) RNA, 17, 2058–2062.
30) Iyer, L.M., Zhang, D., Rogozin, I.B., & Aravind, L. (2011) Nucleic Acids Res., 39, 9473–9497.
31) Boussardon, C., Avon, A., Kindgren, P., Bond, C.S., Challenor, M., Lurin, C., & Small, I. (2014) New Phytol., 203, 1090–1095.
32) Hayes, M.L., Giang, K., Berhane, B., & Mulligan, R.M. (2013) J. Biol. Chem., 288, 36519–36529.
33) Zehrmann, A., Verbitskiy, D., Härtel, B., Brennicke, A., & Takenaka, M. (2010) FEBS Lett., 584, 4181–4186.
34) Wagoner, J.A., Sun, T., Lin, L., & Hanson, M.R. (2015) J. Biol. Chem., 290, 2957–2968.
35) Chung, J.J., Shikano, S., Hanyu, Y., & Li, M. (2002) Trends Cell Biol., 12, 146–150.
36) Tasaki, E., Hattori, M., & Sugita, M. (2010) Plant J., 62, 560–570.
37) Boussardon, C., Salone, V., Avon, A., Berthomé, R., Hammani, K., Okuda, K., Shikanai, T., Small, I., & Lurin, C. (2012) Plant Cell, 24, 3684–3694.
38) Okuda, K. & Shikanai, T. (2012) Nucleic Acids Res., 40, 5052–5064.
39) Okuda, K., Chateigner-Boutin, A.-L., Nakamura, T., Delannoy, E., Sugita, M., Myouga, F., Motohashi, R., Shinozaki, K., Small, I., & Shikanai, T. (2009) Plant Cell, 21, 146–156.
40) Verbitskiy, D., Zehrmann, A., Hartel, B., Brennicke, A., & Takenaka, M. (2012) J. Biol. Chem., 287, 38064–38072.
41) Hayes, M.L., Dang, K.N., Diaz, M.F., & Mulligan, R.M. (2015) J. Biol. Chem., 290, 10136–10142.
42) Bentolila, S., Heller, W.P., Sun, T., Babina, A.M., Friso, G., van Wijk, K.J., & Hanson, M.R. (2012) Proc. Natl. Acad. Sci. USA, 109, E1453–E1461.
43) Zehrmann, A., Härtel, B., Glass, F., Bayer-Császár, E., Obata, T., Meyer, E., Brennicke, A., & Takenaka, M. (2015) J. Biol. Chem., 290, 6445–6456.
44) Glass, F., Härtel, B., Zehrmann, A., Verbitskiy, D., & Takenaka, M. (2015) Mol. Plant, 8, 1466–1477.
45) Hirose, T. & Sugiura, M. (2001) EMBO J., 20, 1144–1152.
46) Tillich, M., Hardel, S.L., Kupsch, C., Armbruster, U., Delannoy, E., Gualberto, J.M., Lehwark, P., Leister, D., Small, I.D., & Schmitz-Linneweber, C. (2009) Proc. Natl. Acad. Sci. USA, 106, 6002–6007.
47) Sun, T., Germain, A., Giloteaux, L., Hammani, K., Barkan, A., Hanson, M.R., & Bentolila, S. (2013) Proc. Natl. Acad. Sci. USA, 110, E1169–E1178.
48) Shi, X., Hanson, M.R., & Bentolila, S. (2015) Nucleic Acids Res., 43, 3814–3825.
49) Shi, X., Germain, A., Hanson, M.R., & Bentolila, S. (2016) Plant Physiol., 170, 294–309.
50) Sun, T., Shi, X., Friso, G., Van Wijk, K., Bentolila, S., & Hanson, M.R. (2015) PLoS Genet., 11, e1005028.
51) Leu, K.-C., Hsieh, M.-H., Wang, H.-J., Hsieh, H.-L., & Jauh, G.-Y. (2016) RNA Biol., May 5: 0. [Epub ahead of print].
52) Zhang, H.-D., Cui, Y.-L., Huang, C., Yin, Q.-Q., Qin, X.-M., Xu, T., He, X.-F., Zhang, Y., Li, Z.-R., & Yang, Z.-N. (2015) Photosynth. Res., 126, 311–321.
53) Faivre-Nitschke, S.E., Grienenberger, J.M., & Gualberto, J.M. (1999) Eur. J. Biochem., 263, 896–903.
54) Chen, M., Herde, M., & Witte, C.-P. (2016) Plant Physiol., pii:pp.02031.2015.
55) Kahlau, S. & Bock, R. (2008) Plant Cell, 20, 856–874.
56) Lu, B., Wilson, R.K., Phreaner, C.G., Mulligan, R.M., & Hanson, M.R. (1996) Mol. Cell. Biol., 16, 1543–1549.

ウルム大学分子植物学(ドイツ),グループリーダー,Heisenberg Fellow. 博士(農学).
1974年鹿児島県に生る.2001年京都大学農学研究科博士課程修了.同年からドイツ,ウルム大学分子植物学研究科にて研究.01~05年博士研究員.05~07年日本学術振興会海外特別研究員.07年よりグループリーダー.11年ドイツHabilitation取得.11年よりDFG, Heisenberg Fellow.
研究テーマと抱負植物オルガネラのRNA編集機構の研究.PPRタンパク質,MORFタンパク質の機能解析.
ウェブサイトhttps://sites.google.com/site/mizukitakenakassite/home
趣味旅行,読書,酒と料理.

This page was created on 2016-08-30T16:33:40.174+09:00
This page was last modified on 2016-10-14T13:09:41.142+09:00
このサイトは(株)国際文献社によって運用されています。