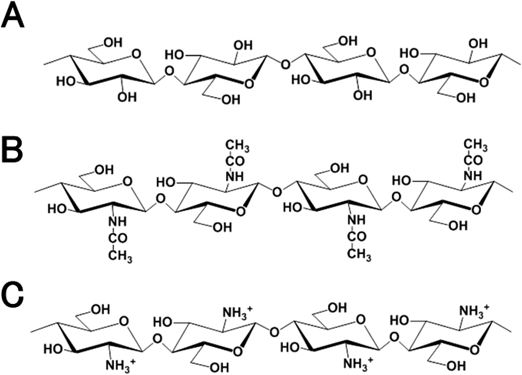
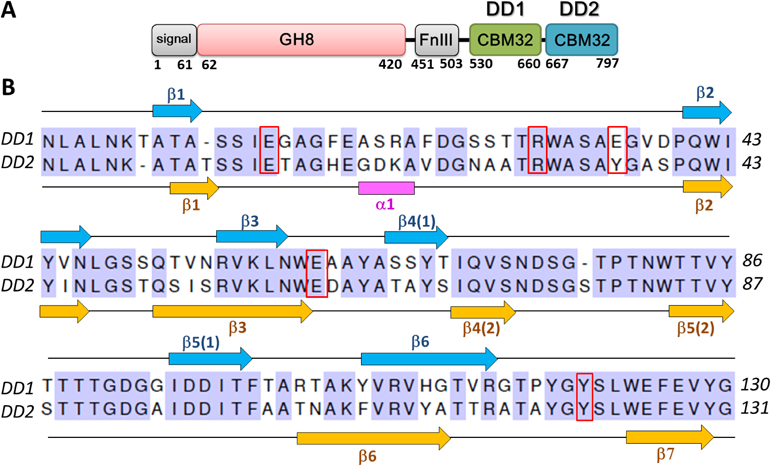
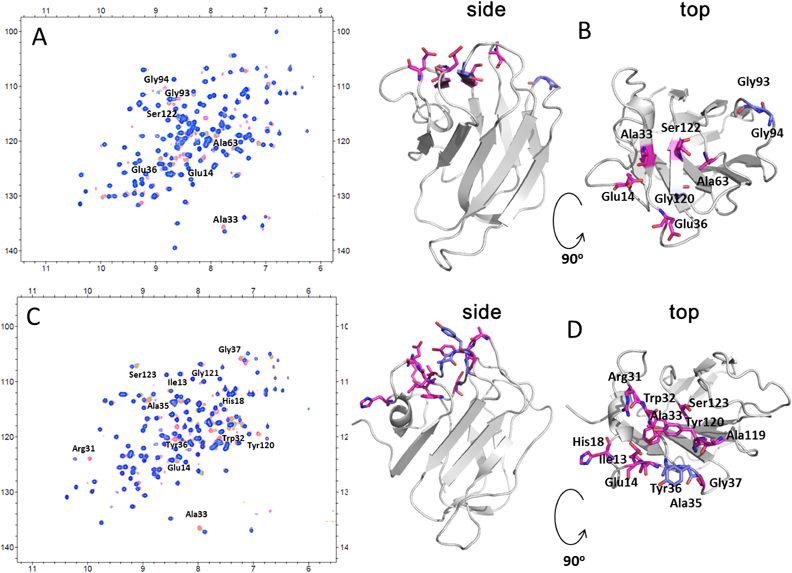
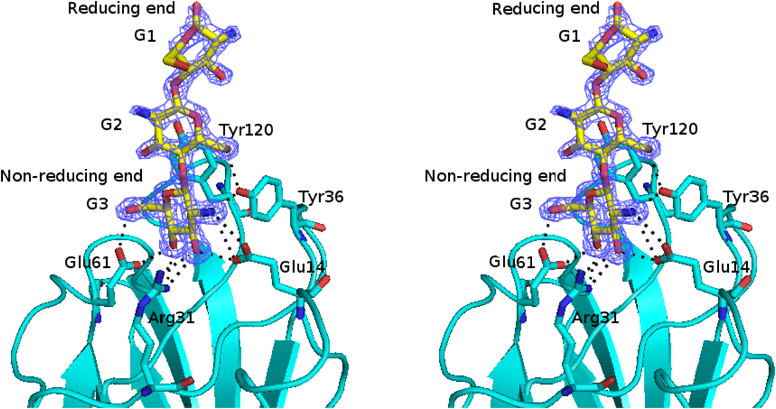
キトサンの尻尾を捕らえるタンパク質Proteins capturing the tail of chitosan
近畿大学大学院農学研究科バイオサイエンス専攻バイオ分子化学研究室Graduate School of Agriculture, Kinki University ◇ 〒631–8505 奈良市中町3327–204 ◇ 3327–204 Nakamachi, Nara 631–8505, Japan
受付日:2016年4月22日Received: April 22, 2016
発行日:2016年10月25日Published: October 25, 2016