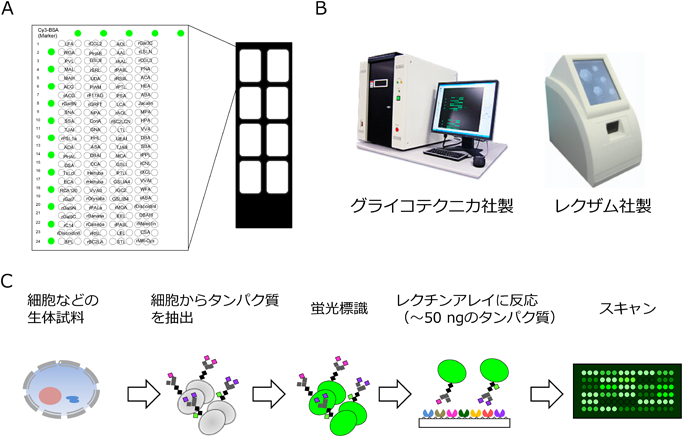
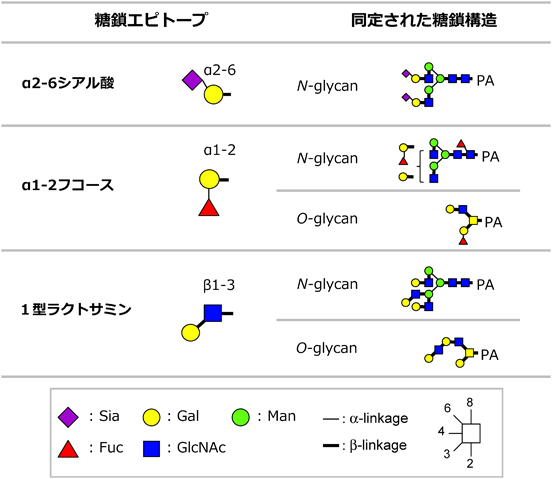
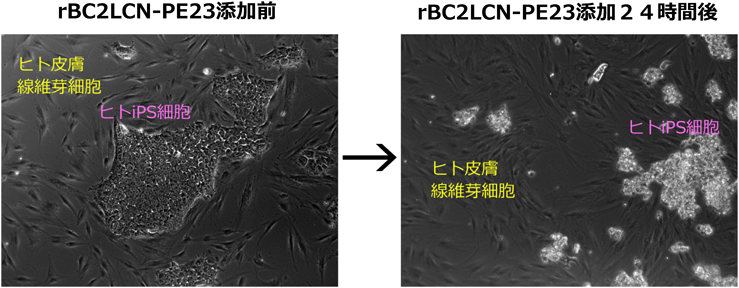
1) Tateno, H., Toyota, M., Saito, S., Onuma, Y., Ito, Y., Hiemori, K., Fukumura, M., Matsushima, A., Nakanishi, M., Ohnuma, K., Akutsu, H., Umezawa, A., Horimoto, K., Hirabayashi, J., & Asashima, M. (2011) J. Biol. Chem., 286, 20345–20353.
2) Hirabayashi, J., Yamada, M., Kuno, A., & Tateno, H. (2013) Chem. Soc. Rev., 42, 4443–4458.
3) Hasehira, K., Tateno, H., Onuma, Y., Ito, Y., Asashima, M., & Hirabayashi, J. (2012) Mol. Cell. Proteomics, 11, 1913–1923.
4) Fujitani, N., Furukawa, J., Araki, K., Fujioka, T., Takegawa, Y., Piao, J., Nishioka, T., Tamura, T., Nikaido, T., Ito, M., Nakamura, Y., & Shinohara, Y. (2013) Proc. Natl. Acad. Sci. USA, 110, 2105–2110.
6) Natunen, S., Satomaa, T., Pitkanen, V., Salo, H., Mikkola, M., Natunen, J., Otonkoski, T., & Valmu, L. (2011) Glycobiology, 21, 1125–1130.
7) Tateno, H., Matsushima, A., Hiemori, K., Onuma, Y., Ito, Y., Hasehira, K., Nishimura, K., Ohtaka, M., Takayasu, S., Nakanishi, M., Ikehara, Y., Ohnuma, K., Chan, T., Toyoda, M., Akutsu, H., Umezawa, A., Asashima, M., & Hirabayashi, J. (2013) Stem Cells Transl. Med., 2, 265–273.
8) Tang, C., Lee, A.S., Volkmer, J.P., Sahoo, D., Nag, D., Mosley, A.R., Inlay, M.A., Ardehali, R., Chavez, S.L., Pera, R.R., Behr, B., Wu, J.C., Weissman, I.L., & Drukker, M. (2011) Nat. Biotechnol., 29, 829–834.
9) Onuma, Y., Tateno, H., Hirabayashi, J., Ito, Y., & Asashima, M. (2013) Biochem. Biophys. Res. Commun., 431, 524–529.
10) Kawabe, K., Tateyama, D., Toyoda, H., Kawasaki, N., Hashii, N., Nakao, H., Matsumoto, S., Nonaka, M., Matsumura, H., Hirose, Y., Morita, A., Katayama, M., Sakuma, M., Kawasaki, N., Furue, M.K., & Kawasaki, T. (2013) Glycobiology, 23, 322–336.
11) Matsumoto, S., Nakao, H., Kawabe, K., Nonaka, M., Toyoda, H., Takishima, Y., Kawabata, K., Yamaguchi, T., Furue, M.K., Taki, T., Okumura, T., Yamazaki, Y., Nakaya, S., Kawasaki, N., & Kawasaki, T. (2015) J. Biol. Chem., 290, 20071–20085.
12) Tateno, H., Saito, S., Hiemori, K., Kiyoi, K., Hasehira, K., Toyoda, M., Onuma, Y., Ito, Y., Akutsu, H., & Hirabayashi, J. (2016) Glycobiology.
13) Hasehira, K., Hirabayashi, J., & Tateno, H. (2016) Glycoconj. J.
14) Wang, Y.C., Stein, J.W., Lynch, C.L., Tran, H.T., Lee, C.Y., Coleman, R., Hatch, A., Antontsev, V.G., Chy, H.S., O’Brien, C.M., Murthy, S.K., Laslett, A.L., Peterson, S.E., & Loring, J.F. (2015) Sci. Rep., 5, 13317.
15) Tateno, H., Onuma, Y., Ito, Y., Minoshima, F., Saito, S., Shimizu, M., Aiki, Y., Asashima, M., & Hirabayashi, J. (2015) Stem Cell Rep., 4, 811–820.
16) Tateno, H., Onuma, Y., Ito, Y., Hiemori, K., Aiki, Y., Shimizu, M., Higuchi, K., Fukuda, M., Warashina, M., Honda, S., Asashima, M., & Hirabayashi, J. (2014) Sci. Rep., 4, 4069.