シュゴシンタンパク質が染色体末端で果たす意外な役割Unexpected roles of Shugoshin at chromosome ends
大阪大学・蛋白質研究所Institute for Protein Research, Osaka University ◇ 〒565–0871 大阪府吹田市山田丘3–2 ◇ 3–2 Yamadaoka, Suita, Osaka 565–0871, Japan
発行日:2017年2月25日Published: February 25, 2017
© 2017 公益社団法人日本生化学会© 2017 The Japanese Biochemical Society
真核生物は,DNAとタンパク質などからなる線状の染色体を持つ.染色体の最末端には,テロメアと呼ばれるドメインが存在する.テロメアは,繰り返し配列(哺乳類の場合,[TTA GGG]n)からなるDNAと,それを基にして集合するさまざまなタンパク質からなる.近年,テロメアに関する研究が飛躍的に進み,テロメアが染色体末端保護に必須であること,永続的な生殖細胞の維持に必須であること,細胞老化のタイミング制御に重要であること,体細胞分裂期や減数分裂期の染色体動態制御に重要であることなどが明らかにされてきた.
テロメアに隣接して,サブテロメアと呼ばれるドメインが存在する.サブテロメアDNAはテロメア繰り返し配列を持たないが,各生物種のサブテロメア間で相同性が高い配列がモザイク状に組み合わさった領域を含む.その共通配列全体の長さは生物種によって異なり,分裂酵母では約50 kb,ヒトでは数百kbに及ぶ.また一般的に,サブテロメアはヒストンH3のK9(9番目のリシン)残基のメチル化を介したヘテロクロマチン構造を有している.
近年,テロメアに関する多くの知見が蓄積してきたのに対して,サブテロメアの機能や制御のメカニズムについては不明な点が多く残されており,サブテロメア研究はまだ黎明期にあるといえる.本稿では,最近我々が発見した,分裂酵母のシュゴシンファミリータンパク質がサブテロメアで果たす意外な役割について紹介する.
分裂酵母は一倍体で安定に生育する単細胞真核生物であり,3本の線状染色体を持つ.1番および2番染色体のテロメアに隣接して約50 kbのサブテロメア共通配列が存在し,その領域を中心としてヘテロクロマチンが形成されている1).ヘテロクロマチンに隣接した約50 kbにわたって,さまざまなヒストン修飾レベルが低く維持されている特殊なクロマチン領域が存在する.この領域は,細胞周期の間期(細胞分裂期以外)にDNA染色剤であるDAPI(4′,6-diamidino-2-phenylindole)で濃染される.このようなサブテロメアの間期特異的な高度に凝縮したクロマチン構造は,knobと呼ばれている2).したがって,分裂酵母のサブテロメアには,ヘテロクロマチンとknobという二つの異なるクロマチン高次構造が隣接して存在していることになる.
一方,3番染色体のテロメア隣接領域の構造は,細胞株によって異なる.両端のテロメアに隣接してribosomal RNA(rRNA)を産生する長大なribosomal DNA(rDNA)繰り返し配列が存在するが,テロメアとrDNA配列の間に,サブテロメア共通配列の一部(約15 kb)を両方のテロメア脇に有する分裂酵母株,片方のテロメア脇に有する分裂酵母株,まったく持たない分裂酵母株がある.よって,分裂酵母のサブテロメア共通配列は,3番染色体のものを含めると,4~6か所存在することになる.
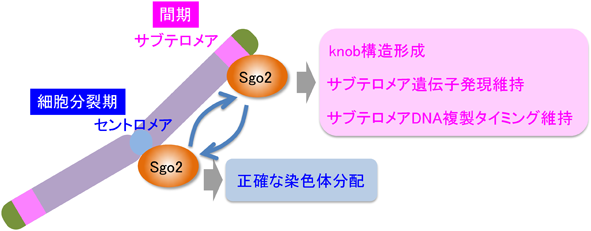
シュゴシンファミリータンパク質は,酵母からヒトまで真核生物に広く保存されており,正確な染色体分配に寄与している.分裂酵母にはシュゴシンタンパク質の二つのパラログが存在しており,Sgo1は減数分裂期特異的に発現してセントロメアに局在し,減数第一分裂期に姉妹染色分体が分かれるのを防ぐ重要な役割を果たす3).一方,Sgo2は恒常的に発現しており,細胞分裂期(M期)にセントロメアに局在し,姉妹染色分体の正確な分配に寄与する4).興味深いことに,Sgo2は間期にテロメア近傍に局在することが報告された4–6).しかし,そのより正確な局在情報やテロメア近傍に局在するSgo2の生理学的役割は不明であった.
そこで我々は,サブテロメアの新規機能・制御メカニズムを明らかにするため,Sgo2のテロメア近傍における機能を探った.まず,間期におけるSgo2の正確な染色体局在をゲノムワイドなChIP-on-chip解析(研究対象タンパク質を認識する抗体を用いてクロマチン免疫沈降を行い,単離したDNAサンプルをゲノムDNAマイクロアレイにハイブリッド形成させ,染色体の各部位の含有量を測定する解析)によって調べたところ,Sgo2は1番および2番染色体のサブテロメア全体(約100 kb)に局在することがわかった.一方,3番染色体においてSgo2の明らかな染色体末端近傍局在は検出されなかった(この研究では3番染色体にサブテロメア共通配列をまったく含まない株を使用した).また,細胞周期を同調させた細胞を用いた実験より,Sgo2のサブテロメア局在は間期の中でG2期においてピークに達することがわかった.
Sgo2のサブテロメア局在はどのように制御されているのだろうか.まず,テロメア構造を失って染色体末端が融合した染色体環状化株においても,Sgo2のサブテロメア局在は失われなかったことから,テロメア構造はSgo2のサブテロメア局在に必要ないことがわかった.さまざまな解析を重ねた結果,Sgo2のサブテロメア局在は,そのセントロメア局在と同様に,Bub1キナーゼ(正確な染色体分配を監視するM期チェックポイント因子)によるヒストンH2Aのリン酸化に大きく依存することが明らかになった.興味深いことに,H2Aのリン酸化は,M期においてはセントロメア領域にほぼ限定されているが,間期においてはサブテロメア以外の領域にも広くみられる6).したがって,H2Aのリン酸化はSgo2のサブテロメア局在にとって必要条件ではあるが,十分条件ではない.現在のところ,間期におけるSgo2の局在をサブテロメアに限局させるファクターについては不明である.
2節で述べたように,分裂酵母細胞では,間期特異的にサブテロメア領域に高度に凝縮したknob構造が形成される.そこで,Sgo2とknobとの関係を探った.驚いたことに,Sgo2を欠損させるとknobがまったく観察されなくなった.同様に,Bub1欠損株でもknobがまったくみられなかったことから,Sgo2がサブテロメアに局在することがknob形成に必須であることがわかった.
一方,Sgo2の欠損は,テロメア隣接領域に形成されるヘテロクロマチンには影響を及ぼさなかった.逆に,ヘテロクロマチンを破壊しても,Sgo2の局在は大きく変化しなかった.これらのことから,Sgo2はサブテロメアヘテロクロマチン形成には必要ではなく,Sgo2とヘテロクロマチンは基本的に独立であると考えられた.
一般的に,高度に凝縮したクロマチン構造は,その領域に存在する遺伝子発現を抑制する効果をもたらすことが知られている.そこで,Sgo2がサブテロメア領域の遺伝子発現維持に関与しているかどうか探ったところ,Sgo2欠損株ではサブテロメア領域に存在する遺伝子の転写が顕著に増加していることがわかった.さらに,RNAポリメラーゼIIの局在や,転写に関連するヒストン修飾(ヒストンH3-K4やH3-K36のメチル化)もサブテロメア領域において顕著に増加していた.Sgo2欠損によるこれらの影響は,セントロメアにおいては一切みられなかったことから,Sgo2はサブテロメア特異的に転写過程を抑制していると考えられた.
細胞周期のS期(DNA合成期)におけるDNA複製は,染色体全体で一斉に起こるのではなく,部位によって複製のタイミングが決まっている.分裂酵母のサブテロメアには,S期の遅い時期にDNA複製が開始される複製開始点(late origin)が集中して存在する7).最近,テロメア結合タンパク質がテロメア近傍だけでなく,染色体の広い範囲におけるlate originの複製タイミング維持に寄与していることが報告されているが8, 9),まだ全容は明らかになっていない.そこで,Sgo2欠損株でサブテロメアの複製タイミングを解析したところ,Sgo2が局在するサブテロメア領域のlate originにおいてのみ時期尚早なDNA複製が起きていた.これはDNA複製開始因子であるSld3のoriginへのリクルートの抑制が解除されていることが原因であった.以上のことから,Sgo2はSld3のlate originへのリクルートを抑えることによってサブテロメア領域のDNA複製タイミングを維持していることが明らかになった.
分裂酵母のシュゴシンタンパク質Sgo2は,M期にはセントロメアに局在して正確な染色体分配に寄与していることが知られていたが,間期になるとサブテロメア全体に局在し,サブテロメアのknob構造形成,遺伝子発現維持,DNA複製タイミング維持に重要な役割を果たしていることが明らかになった(図1)10).セントロメアタンパク質として有名なシュゴシンが,サブテロメアでセントロメアでの機能とは別種の働きをしているというのは驚きである.
ここで多くの疑問が浮上してくる.たとえば,Sgo2は具体的にどのようにしてサブテロメアで機能を発揮しているのだろうか? 中でも興味深いのは,Sgo2がどのようにしてknob構造形成に関わっているのか,Sgo2はセントロメアではなくサブテロメアでのみknob構造形成を誘導するのはなぜか,である.M期のシュゴシンは,脱リン酸化酵素の一種であるPP2A(protein phosphatase 2A)と相互作用することによって標的タンパク質のリン酸化レベルを制御していることが報告されている11).間期のSgo2も脱リン酸化酵素をリクルートし,サブテロメアのクロマチンタンパク質を脱リン酸化することによってknob構造形成を誘導しているのかもしれない.ここで,セントロメアとサブテロメア,あるいはM期と間期の違いが鍵になると思われる.
また,Sgo2はknob構造を介してサブテロメアの遺伝子発現やDNA複製タイミングを制御しているのか,あるいはSgo2自身がknob構造とは関係なく制御しているのか,という点も重要である.前者の場合,knobという高度の凝縮したクロマチン構造によってさまざまなタンパク質(RNAポリメラーゼIIやDNA複製開始因子など)がサブテロメアにアクセスできなくなっているという可能性が考えられる.一方,DNA複製の開始には,DDK(Dbf4-dependent kinase)による複製開始点局在タンパク質群のリン酸化が必要とされることが知られている.テロメア結合タンパク質の一つであるRif1はPP1(protein phosphatase 1)を複製開始点にリクルートしてDDKによるリン酸化を打ち消すことによってlate originのDNA複製タイミングを制御していると考えられている12).同様に,Sgo2もPP1あるいはPP2AをサブテロメアにリクルートすることによってDNA複製タイミングを制御しているのかもしれない.
ヒトでは,サブテロメア構造の異常による疾患が知られている.サブテロメアのヘテロクロマチン構造の異常によって筋ジストロフィーを発症することがある.また,サブテロメアDNAの微細な欠失や重複によってサブテロメア微細構造異常症という総称で知られる疾患になることがある.いずれもサブテロメアに存在する遺伝子の発現量の異常が直接的原因と考えられている.現在のところ,分裂酵母以外の生物種のサブテロメアにおいても,ヘテロクロマチン以外にknobのような高度の凝縮したクロマチン構造が存在するのか,シュゴシンタンパク質がサブテロメアに局在するのかは不明である.しかし,サブテロメア遺伝子の発現量の維持がヒトの健康にきわめて重要であることから,何らかの制御機構が存在していることが推測される.
本稿の元となった論文の共著者の皆様に感謝いたします.
1) Kanoh, J., Sadaie, M., Urano, T., & Ishikawa, F. (2005) Curr. Biol., 15, 1808–1819.
2) Matsuda, A., Chikashige, Y., Ding, D.Q., Ohtsuki, C., Mori, C., Asakawa, H., Kimura, H., Haraguchi, T., & Hiraoka, Y. (2015) Nat. Commun., 6, 7753.
3) Kitajima, T.S., Kawashima, S.A., & Watanabe, Y. (2004) Nature, 427, 510–517.
4) Kawashima, S.A., Tsukahara, T., Langegger, M., Hauf, S., Kitajima, T.S., & Watanabe, Y. (2007) Genes Dev., 21, 420–435.
5) Vanoosthuyse, V., Prykhozhij, S., & Hardwick, K.G. (2007) Mol. Biol. Cell, 18, 1657–1669.
6) Kawashima, S.A., Yamagishi, Y., Honda, T., Ishiguro, K., & Watanabe, Y. (2010) Science, 327, 172–177.
7) Hayashi, M., Katou, Y., Itoh, T., Tazumi, A., Yamada, Y., Takahashi, T., Nakagawa, T., Shirahige, K., & Masukata, H. (2007) EMBO J., 26, 1327–1339.
8) Hayano, M., Kanoh, Y., Matsumoto, S., Renard-Guillet, C., Shirahige, K., & Masai, H. (2012) Genes Dev., 26, 137–150.
9) Tazumi, A., Fukuura, M., Nakato, R., Kishimoto, A., Takenaka, T., Ogawa, S., Song, J.H., Takahashi, T.S., Nakagawa, T., Shirahige, K., & Masukata, H. (2012) Genes Dev., 26, 2050–2062.
10) Tashiro, S., Handa, T., Matsuda, A., Ban, T., Takigawa, T., Miyasato, K., Ishii, K., Kugou, K., Ohta, K., Hiraoka, Y., Masukata, H., & Kanoh, J. (2016) Nat. Commun., 7, 10393.
11) Kitajima, T.S., Sakuno, T., Ishiguro, K., Iemura, S., Natsume, T., Kawashima, S.A., & Watanabe, Y. (2006) Nature, 441, 46–52.
12) Davé, A., Cooley, C., Garg, M., & Bianchi, A. (2014) Cell Reports, 7, 53–61.

大阪大学蛋白質研究所細胞核ネットワーク研究室独立准教授.理学博士(東京大学).
1991年東京大学理学部生物化学科卒業.96年同大学院理学系研究科生物化学専攻博士課程修了.東京大学医科学研究所ポスドク研究員,米国Scripps研究所ポスドク研究員などを経て,2000年東京工業大学生命理工学研究科助手.02年京都大学生命科学研究科助教.09年大阪大学蛋白質研究所テニュアトラック准教授.13年より現職.
研究テーマと抱負真核生物の線状染色体末端領域のテロメアとサブテロメアのクロマチン構造の機能や制御メカニズムを研究している.今後は,ヒトや大型類人猿細胞で研究を展開し,サブテロメアと進化や疾患との関係も探っていきたい.
ウェブサイトhttp://www.protein.osaka-u.ac.jp/icr/network/
趣味フィギュアスケート(観る),音楽(聴く,奏でる,歌う),写真,料理.

This page was created on 2016-12-21T14:29:03.289+09:00
This page was last modified on 2017-02-17T18:26:09.381+09:00
このサイトは(株)国際文献社によって運用されています。