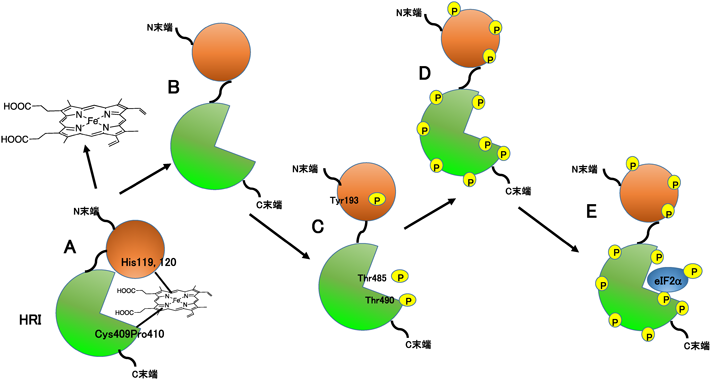
2. 3価ヘム鉄の結合・解離が機能を制御するヘムセンサーHRI
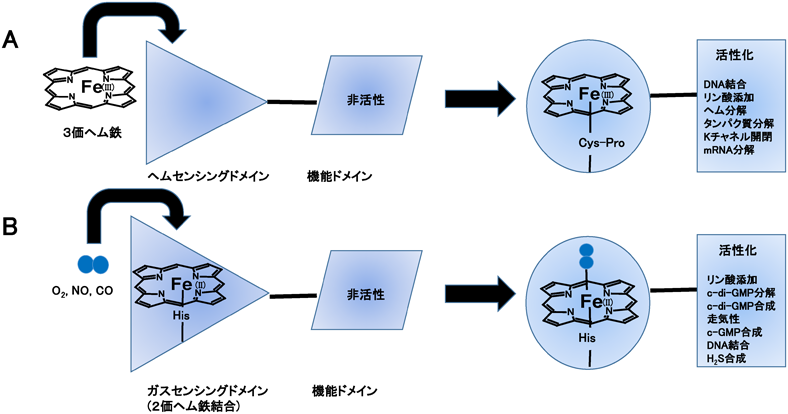
ヘムセンサーでは,ヘム鉄自身,特に3価へム鉄が第一シグナルとして機能性タンパク質と結合・解離を行い,その結果,ヘム鉄結合部位周辺のタンパク質構造が変化する.この構造変化が第二シグナルになり,DNA結合やタンパク質リン酸化(キナーゼ)反応などのスイッチの切り替え(on/off)を制御する9–13)(図1A).我々は,HRI[heme-regulated inhibitor,ヘム制御真核生物翻訳開始因子2αキナーゼ(heme-regulated eukaryotic initiation factor 2α kinase)ともいう]の全長タンパク質の大腸菌での発現系を構築し,各種変異体の活性やスペクトルを検討し,3価ヘム鉄の結合・解離で制御されるキナーゼ反応の分子機能の一部を明らかにした.
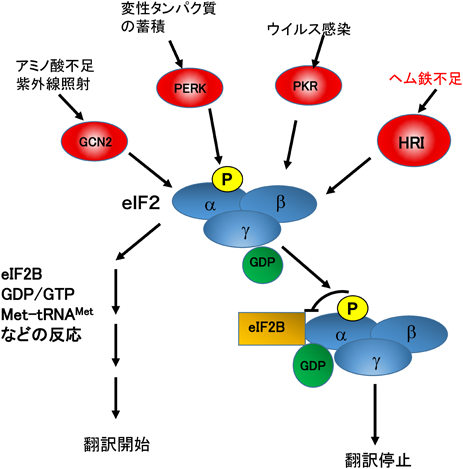
HRIはストレスに対応してタンパク質の翻訳・合成を初期段階で停止させる四つのキナーゼ(GCN2, PERK, PKR, HRI)の一つである.タンパク質の翻訳・合成初期過程はα, β, γサブユニットから構成される真核生物翻訳開始因子2(eukaryotic initiation factor 2:eIF2)が関わるいくつかの複雑な反応で構成されている14)(図2).さまざまなストレス,たとえば,(1)紫外線照射,アミノ酸不足,(2)変性タンパク質の蓄積(ERストレス),および(3)ウイルス感染,を細胞が感知すると,それぞれのストレスに対応して,GCN2, PERK, PKRというキナーゼが活性化する.その結果,共通の基質eIF2のαサブユニット(eIF2α)の51番目のセリン残基がリン酸化され,タンパク質の翻訳・合成は初期段階で停止する.HRIは,これらの三つのキナーゼと同じ活性部位と基質を保持し,赤血球内で3価ヘム鉄の不足というストレスを感知し,グロビンの翻訳・合成をその初期段階で停止させる9–11, 14, 15)(図2).
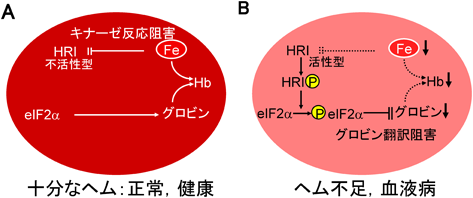
赤血球内では,ヘム鉄とグロビンタンパク質が1:1で結合してヘモグロビンが生成されており,ヘム鉄とグロビンタンパク質の適切な濃度均衡維持が必要となる.ヘム鉄濃度に比例したグロビン濃度の調節は転写段階の制御によるのではない15)(図3).まず,赤血球内の3価ヘム鉄濃度が正常である場合には,HRIタンパク質に3価ヘム鉄が結合することによってHRI活性が抑制されている(図3).ところが,ヘム鉄が正常に生合成されないようなある種の血液疾患においては,赤血球内の3価ヘム鉄の濃度が低下する.すると3価ヘム鉄がHRIから解離してHRIの大規模なタンパク質の構造変化が誘起され,キナーゼ活性部位がタンパク質分子の表面に露出する.それによってHRIはまず自己リン酸化して活性型になり,最終的にeIF2αに対するキナーゼ活性を発現する.eIF2αがリン酸化されると,タンパク質翻訳・合成は初期段階で停止する.こうして低濃度の3価ヘム鉄に応答したグロビンタンパク質の合成抑制が達成される15).
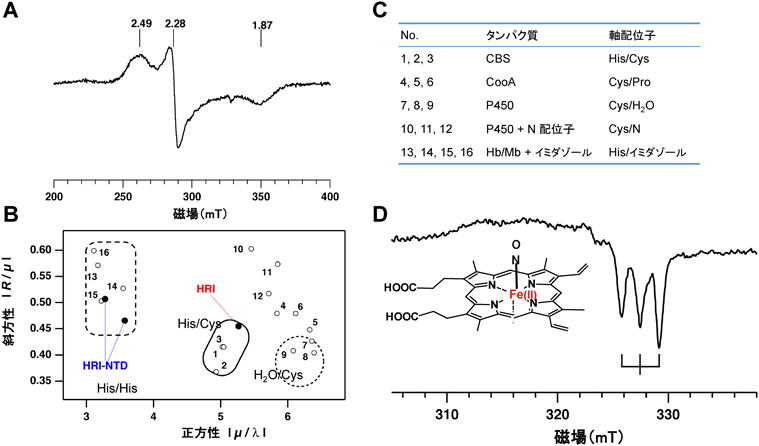
一般に,ヘムセンサーの3価ヘム鉄のセンシング部位は,システイン(Cys),またはシステイン–プロリン(Cys–Pro)配列を有し,この配列はCPモチーフ,またはヘム制御モチーフ(heme regulatory motif:HRM)とも呼ばれる16).3価ヘム鉄が結合したHRI全長タンパク質の吸収スペクトルは423 nmに吸収極大を示し,軸配位子が上下から3価ヘム鉄に結合した6配位低スピン錯体であると推定された17).また,その電子常磁性共鳴(EPR)スペクトルの結晶場ダイアグラム(crystal field diagram)のパラメーターは,6配位ヒスチジン–3価鉄–システイン(His–Fe(III)–Cys)の配位構造をとるヘムタンパク質の領域に存在した(図4A~C).
さらに,N末端領域をさまざまの長さで欠損させた変異体を作製し,それらの吸収スペクトルを全長タンパク質と比較すると,N末端85残基を欠損させた変異体では野生型と同様に423 nm付近に6配位His–Fe(III)–Cys配位構造に由来するソーレー帯の吸収極大を示した.しかし,N末端127残基,あるいは145残基を欠損させた変異体では370 nm付近に5配位Fe(III)–Cys配位構造に由来する吸収極大を示し,EPRスペクトルには5配位Fe(III)配位構造に由来するシグナルが検出された.これらのことより,N末端127残基までに存在するヒスチジン残基,および128番目のアミノ酸残基以降の領域に存在するシステイン残基が野生型HRIの6配位His–Fe(III)–Cysの軸配位子であることが推定された18).
ヘムの軸配位子であるアミノ酸残基を同定するためにHRIのN末端に存在する7個のヒスチジン残基の部位特異的変異体(H75L, H78L, H80A, H86A, H119A, H120A, H126A)を構築しそのヘム結合性を調べた.しかし,これらの一重置換変異体は野生型と同様のヘム結合性を示し,唯一H119A/H120Aの二重変異体でヘム結合性の消失が認められた.この現象は,HRIタンパク質が柔軟性を有しており,H119AまたはH120Aの一残基置換では,隣接するヒスチジン残基と配位子交換反応が進行するためにヘム結合性は変わらず,交換反応が起こらない二重変異体にすることでヘム結合性の消失が観測できたと解釈され,His119またはHis120が3価ヘム鉄の軸配位子の一つであると推定された19).次に,C末端側にある6個のシステインの部位特異的変異体(C208S, C385S, C409S, C464S, C491S, C550S)を構築し,そのヘム結合性を調べた.その結果,C409S変異体では3価ヘム鉄への結合性が消失しており,Cys409が3価ヘム鉄の軸配位子であることが示唆された19)(図5).このCys409はC末端側に二つ存在するCPモチーフの一つであったことから,隣接するプロリン残基Pro410をアラニン置換したP410Aを作製したところ,この変異体も3価ヘム鉄への結合性が消失していた19).このことより,Pro410は,Cys409の3価ヘム鉄との相互作用を立体的に助ける役割をすることが示された.
ここで注意しなければならないことがある.それは,CPモチーフがあくまで3価ヘム鉄の結合部位であるということである,すなわち,3価ヘム鉄が2価ヘム鉄へ還元されると,軸配位子であるシステイン残基が2価ヘム鉄から解離し,この残基に代わって同じタンパク質内の近隣に存在する他のアミノ酸残基(イミダゾール側鎖を有するヒスチジン残基など)が非特異的に2価ヘム鉄に結合・配位して,ヘム鉄結合部位周辺のタンパク質構造が著しく変化する9–11).一方,酸素やCOは2価ヘム鉄にしか結合しない.3価ヘム鉄しか結合しない(感知する)ヘムセンサータンパク質が,同時に2価鉄でないと機能できないヘム基盤COセンサーになるといういくつかの報告があるが,還元電位より推定されるヘム鉄の還元のしやすさなども考慮する必要がある.
ヘムセンサーでは,3価ヘム鉄のタンパク質への親和性はヘモグロビンやミオグロビンより著しく低い.3価ヘム鉄のHRIタンパク質からの解離速度定数は1.5×10−3 s−1でミオグロビンの値8.4×10−7 s−1より103倍以上高い9, 18).そのために,3価ヘム鉄はHRIタンパク質に対して容易に結合・解離し,タンパク質の構造変化を誘起してその機能を発揮すると推定される.また,赤血球内の遊離3価ヘム鉄の濃度と3価ヘム鉄のHRIからの解離平衡定数とは近い値であると推定される.一方,核内で3価ヘム鉄が結合する転写制御因子Rev-erbβの3価ヘム鉄の解離速度定数は10−6 s−1で非常に低い20).核内の3価ヘム鉄の濃度は最大2.5 nM程度ときわめて低いため13),核内で3価ヘム鉄の結合・解離が転写やDNAとの結合を制御するヘムセンサーは3価ヘム鉄との親和性がきわめて高く,その解離平衡定数はヘム鉄の核内濃度pM~nM程度と推定される.
HRIでは,3価ヘム鉄の二つの軸配位子であるHis119とCys409は一次構造上かなり離れている19).そこで,His119を含むN末端側とCys409を含むC末端側をドメインごとに過剰発現し,両ドメイン間のタンパク質–タンパク質相互作用が3価ヘム鉄によってどう影響されるかをプルダウンアッセイで調べた.各ドメインタンパク質は,3価ヘム鉄がある場合にのみタンパク質–タンパク質相互作用することが確認できたが,3価ヘム鉄がない場合には相互作用は確認できなかった.また,3価ヘム鉄の軸配位子であるH119A/H120Aの二重変異体やC409S変異体では,3価ヘム鉄存在下においてもタンパク質–タンパク質相互作用が観察されなかった.これらの結果から,HRIタンパク質において,3価ヘム鉄の結合・解離に伴う,閉鎖型(Closed form)/開放型(Open form)という大きな構造変化が誘起されることが推定された9, 19, 21).Bach1/2やイオンチャネル(下記参照)においても,ヘム結合部位周辺のタンパク質構造の高い柔軟性が示唆されている22, 23).
NOはさまざまな重要な生理機能を制御している.3価ヘム鉄が結合した不活性型HRIにNOを添加すると,3価ヘム鉄がNOによって2価ヘム鉄へ還元されると同時に5配位Fe(II)–NO錯体へ変化し,HRIは活性化される17).その分子機構については,5配位2価ヘム鉄–NO錯体になると,ヘム鉄とHRIタンパク質間の結合(配位)が解消されてヘム鉄がタンパク質から離れて浮いた状態になるために(図4Dの配位構造参照),HRIのキナーゼ活性部位が開放されると推定される.また,N末端85残基を欠損したHRI変異体では,NO添加により6配位NO–Fe(II)–Hisが生成し,野生型酵素で観察されたようなNOによる活性回復が観測されなかった.この結果から,活性型5配位NO–Fe(II)錯体の生成にはHRIのN末端領域が必要であると推定された17, 18).
HRIの自己リン酸化とヘム鉄の結合性の関連を調べたところ,HRIで自己リン酸化する33個のアミノ酸が質量分析で同定された.これら自己リン酸化部位の部位特異的変異体のキナーゼ活性を調べると,自己リン酸化はまずTyr193, Thr485,およびThr490で起こり,続いて残り30個のアミノ酸(Ser, Thr, Tyr)がリン酸化した後にHRIが活性型になり,その後に基質であるeIF2αのSer21がリン酸化されることが示唆された24)(図5).また,HRIがいったんリン酸化すると3価ヘム鉄のHRIへの結合が妨げられることから,リン酸化がタンパク質表面の修飾のみならず,タンパク質全体構造の変化を伴うことが示唆された.
水銀はチオールに対して高い親和性がある.そのため,当然ながら,システインが軸配位子である3価ヘム鉄結合型全長HRIへ水銀を添加することによりシステイン軸配位は消失した.一方,3価ヘム鉄が結合していない活性型HRIに水銀を添加すると,強い活性阻害(IC50=0.6 μM)が認められ,この活性阻害はNOの添加により消失した.このことから,キナーゼ活性にはシステインが関わっている可能性が示唆された.また,NOはチオール基とSNOを形成することから,水銀によりブロックされたシステインの機能がNOによって回復したと推定される25).
3価ヘム鉄は,GDCR8のシステイン残基に結合してマイクロRNAのプロセッシングに寄与する他26–28)HAP116),ALAS29),Bach1/222, 30–34),p5335),Shu1/Fep136),PpsR37),All497838),SbnI39),NPAS240–46),Per147),Per248–51),Rev-erbβ20, 52, 53),E7553–55)のシステイン残基への結合・解離を介して,標的タンパク質遺伝子の転写制御やDNA結合などの機能制御に関与することが報告されている.先述したように,これらの反応はヘム鉄の濃度が低い核内で進行することから,3価ヘム鉄が標的タンパク質に対して非常に高い親和性を有すると推定される.NPAS2については,3価ヘム鉄以外に,2価ヘム鉄,pH, NADPHがDNA結合や他の転写制御タンパク質との相互作用に関係しているという報告もある44–46).
膜結合性イオンチャネルでは3価ヘム鉄がシステイン残基へ結合して,チャンネルの開閉を制御する23, 56–60).IRP261)やIrr62–64)では,3価ヘム鉄のシステイン残基への結合・解離が,タンパク質分解やヘムの分解を制御する.
ヘムセンサーでもヘム結合(センシング)部位がシステイン残基ではなくヒスチジン残基の場合もある.ヘム分解に関わるHutZ65),代謝や体内時計に関わるRev-erbα66–68),転写に関わるHrtR69),およびtRNA合成に関わるTrpRS70)では3価ヘム鉄はシステイン残基ではなくヒスチジン残基に結合する.二成分シグナル伝達(下記参照,図8)に関わるヘムセンサー・キナーゼChrSでは3価ヘム鉄の結合部位は同定されていない71, 72).
ヘム酸素添加酵素HO-2にはCPモチーフが備わっており,その部位への3価ヘム鉄の結合が研究されているが,ヘム鉄の結合がHO-2の機能に果たす役割については不明である73, 74).
PGRMC1(progesterone-receptor membrane component 1)は3価ヘム鉄がチロシン残基に結合・解離して上皮成長因子受容体やシトクロムP450とのタンパク質相互作用を制御するヘムセンサーであると報告された75).2分子の3価ヘム鉄はスタッキングして直接結合し,Tyr113のみが軸配位子である5配位結合になっている.ヘム鉄のスタッキング二量体とチロシン軸配位結合は,ヘム輸送タンパク質にみられる76, 77).多くの3価ヘム鉄輸送タンパク質のヘム鉄結合部位(軸配位子)もチロシン残基である76–84).PGRMC1はヘム鉄輸送タンパク質であるとも報告されている85).PGRMC1のヘム鉄の還元電位は−331 mVと非常に低い86).ヘム鉄輸送タンパク質での3価ヘム鉄へのチロシン配位は,3価ヘム鉄を安定化し,2価ヘム鉄への還元を難しくしているためではないかと推定される.
酸素は呼吸や代謝などできわめて重要な働きをする.ヘム基盤酸素センサーはN末端の酸素センシングドメインとC末端の機能ドメインから構成される(図1B).酸素分子自身が第一シグナルになり,酸素センシングドメインに存在する2価ヘム鉄に結合・解離してヘム鉄周辺のタンパク質構造変化を誘起し,その構造変化が第二のシグナルになり,C末端側のドメインでの触媒反応を制御する12, 90, 91).
1)ヘム結合PAS構造ドメインを持つ酸素センサー
PAS[Per(Drosophila period clock protein),Arnt(vertebrate aryl hydrocarbon receptor nuclear translocator),and Sim(Drosophila single-minded protein)]ドメインは主にβシートから構成され,多くの分子間,および分子内シグナル伝達に関わっている92).多くのヘム基盤ガスセンサーでは,N末端にあるPASドメインにヘム鉄が結合して,ガスセンシングドメインになる.酸素などのガス分子がこのヘム鉄へ結合することにより,PASタンパク質の構造が著しく変化する93–95).
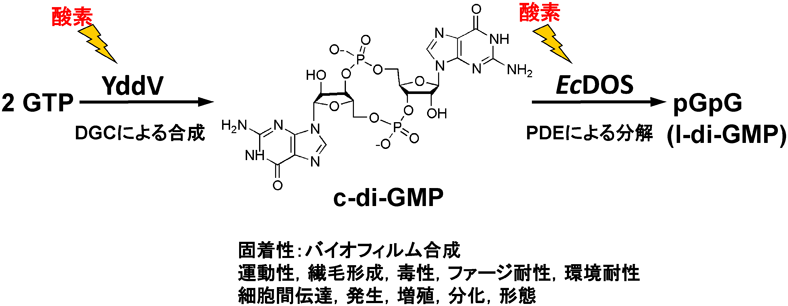
大腸菌由来EcDOS (EcDosP)はヘム結合PASドメインを保持するヘム基盤酸素センサーである.酸素がEcDOSの酸素センシングドメインの2価ヘム鉄へ結合することにより,センシングドメインタンパク質の大きな構造変化が起きる94, 95).そのタンパク質の構造変化がシグナルになって,C末端側の機能ドメインに伝達される.その結果,微生物にとって重要なシグナル分子であるc-di-GMP96)がpGpG(またはlinear-di-GMP)へ分解・変換される反応(phosphodiesterase:PDEによる)が,9.7 min−1から61 min−1へと亢進する(図6)97–100).この反応が機能するC末端側の機能ドメインにはPDEに関わるEALサブドメインとc-di-GMPの合成(diguanylate cyclase:DGCによる)に関わるGGDEFサブドメインの両方のサブドメインを保持している.しかし,EcDOSではPDE活性しか発現しない.大腸菌では,次項で述べるようにDGC活性を保持する別のヘム基盤酸素センサー,YddVが存在し,細胞内の酸素濃度に依存してc-di-GMPの合成や分解がEcDOSとYddVによって相乗的に行われてc-di-GMPの細胞内濃度を部分的に調節していると推定される(図6).
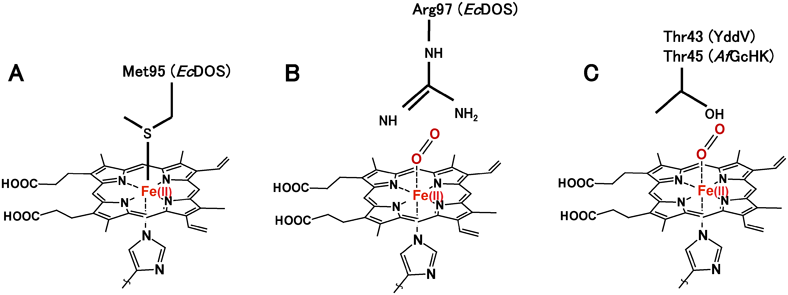
酸素が結合していない不活性型EcDOSでは,Met95が2価ヘム鉄への軸配位子である(図7A)94).2価ヘム鉄に酸素が結合すると,ヘム鉄遠位側(酸素結合側,distal side)のタンパク質の大きな構造変化が誘起され,その結果Arg97がO2–Fe(II)–His錯体の酸素と総合作用するようになる(図7B)95).この酸素の結合に誘起されたタンパク質の構造変化がシグナルになり,そのシグナルがC末端ドメインに伝達され,C末端ドメインにあるc-di-GMP分解触媒(PDE)活性が促進される(図1B,図6)100).
Met95をアラニン,ロイシンなどの2価ヘム鉄に配位しないアミノ酸に変異する(M95A, M95L)と,それらの変異体では酸素の結合がない状態でもすでに活性型であった.M95A,およびM95L変異体の場合,遠位側のアミノ酸Ala95, Leu95がすでに2価ヘム鉄から上位へ解離している.そのために,これら変異体では遠位側のタンパク質構造は酸素が結合したのと類似した構造になっており,酸素が結合しなくても活性体であると推定される.しかし,Met95をヒスチジン残基に変異すると(M95H),野生型と同様にヒスチジン残基は2価ヘム鉄の軸配位子になり,酸素が存在しない状態では不活性型で,酸素を添加することにより活性化する.以上のことより,酸素が2価ヘム鉄へ結合することによりMet95はヘム鉄より解離し,元々遠位に位置していたMet95を含む領域のポリペプチド鎖が上へ押し上げられて,その結果センシングドメイン表面の構造が変化し,その変化がC末端側の活性ドメインに伝わり活性が発現すると推定される100).別の言葉でいうと,Met95は反応をロックするために備わっており,酸素の結合がそのロックを解き放すとも解釈できる.さらに,興味あることに,ヘム鉄が結合していない変異体が,すでに酸素活性型のコンホメーションをとっていることから,ヘム鉄の結合自体が反応をロックしているとも考えられる101).
Met95やArg97の部位特異的変異体では,酸素の結合・解離,自動酸化速度,酸化還元電位などの値102–105)や,共鳴ラマンスペクトルのパラメーター106–108)は野生型EcDOSより著しく異なる(表1,表2).たとえば,M95A, M95L変異体では,酸素が2価ヘム鉄に結合する際にMet95が2価ヘム鉄から解離する必要がないため酸素に対する結合速度定数は野生型EcDOSの30倍以上高くなり,また,解離平衡定数は野生型EcDOSの5%以下になり,酸素に対する親和性は著しく高くなる(表1).また,これらの変異体では,還元電位も野生型EcDOSに比べて著しくマイナス側へ低下する(70~190 mV vs. SHE)(表2)105).したがって,Met95の配位は2価ヘム鉄の安定性に寄与しており,2価ヘム鉄が3価ヘム鉄への酸化を妨げる役目もしていると推定される.
表1 主な酸素センサー,それらの部位特異的変異体,およびミオグロビンへの酸素の結合速度定数kon,解離速度定数koff,解離平衡定数Kd,および自動酸化速度定数kox12, 91, 98, 102, 103, 121, 128) | kon (μM−1s−1) | koff (s−1) | Kd (μM) | kox (min−1) |
|---|
| EcDOS野生型 | 0.0019~0.031 | 0.61~0.64 | 20~340 | 0.0058 |
| Met95Ala | >1 | 0.73 | <0.73 | 0.0013 |
| Met95His | >1 | 0.79 | <0.79 | 0.018 |
| Met95Leu | >1 | 0.45 | <0.45 | 0.0017 |
| Arg97Ala | 0.076 | | | >9.5 |
| Arg97Glu | | | | >46 |
| Arg97Ile | 0.16 | 78 | 500 | 0.16 |
| BjFixL | 0.14 | 20 | 140 | 0.045 |
| RmFixLH | 0.22 | 6.8 | 31 | 0.022 |
| YddV野生型 | 0.9~1.4 | 13 | 9.3~14 | 0.0092 |
| Tyr43Ala | 酸素は結合しない | | | |
| Tyr43Leu | 酸素は結合しない | | | |
| Tyr43Phe | 4,6 | 53 | 12 | 0.069 |
| Tyr43Trp | 4,6 | >150 | >33 | 0.12 |
| AfGcHK野生型 | 1.3, 0.15 | 0.10 | 0.077, 0.67 | ≪0.001 |
| Tyr45Phe | 1.8, 0.15 | 0.35 | 0.19, 0.23 | ≪0.001 |
| Tyr45Leu | 1.5, 0.078 | 8.0 | 5.3, 100 | ≪0.001 |
| Tyr45Trp | 1.3, 0.12 | 3.4 | 2.6, 28 | ≪0.001 |
| SW Mb | 17 | 15 | 0.91 | 0.001 |
| SW Mb:マッコウクジラミオグロビン.Kd(=kon/koff)値が低いほど酸素との親和性が高い. |
表2 主な酸素センサー,その変異体,NOセンサー,ヘムセンサー,COセンサーとヘムタンパク質の還元電位12, 86, 98, 102, 105, 109, 121, 150, 181) | 還元電位(mV vs. SHE) |
|---|
| EcDOS野生型 | +24~66 |
| Met95Ala | −26 |
| Met95His | −122 |
| Met95Ile | −1 |
| Arg97Ala | +43 |
| Arg97Glu | +40 |
| Arg97Ile | +49 |
| BjFixL | +68 |
| ReFixL | +19 |
| YddV | −22 |
| ウシsGC | +187 |
| PGRMC1 | −331 |
| CBS | −350 |
| Aplysia Mb (pH 6.0) | +137 |
| SW Mb | +55 |
| 西洋わさびペルオキシダーゼ | −250 |
| シトクロムP450cam | −300 |
| シトクロムb562 | +113 |
| シトクロムc | +260 |
| Aplysia Mb:アメフラシミオグロビン.SW Mb:マッコウクジラミオグロビン.一般に,還元電位が+だと2価ヘム鉄が安定,−だと3価ヘム鉄が安定に存在する. |
Arg97はO2–Fe(II)–His錯体の酸素と相互作用をして,EcDOSの酸素結合体を安定化する(図7B).この残基をアラニンやグルタミン酸へ置換した変異体では,自動酸化速度が野生型EcDOSより2000倍以上上昇した102)(表1).
EcDOSの各種変異体の共鳴ラマンスペクトルの結果より,Trp53, Met95, Arg97, Tyr126は,ヘム鉄のプロピオン酸やビニル基と水素結合ネットワークを形成していると示唆された.酸素などの軸配位子が2価ヘム鉄に結合することによりこれらの水素ネットワークが変化し,その情報がC末端ドメインに伝達されて活性が制御されると推定された106–108).
ヘム結合PAS構造ドメインを持つ酸素センサーであるEcDOSやFixLの酸素の解離解離定数Kdは20~340 μMと酸素の親和性は比較的低い12, 91, 98).ゆえに,これらの酸素センサーは酸素濃度の比較的高い環境で機能し,極微小の酸素濃度の変化を感知して,その機能を発揮すると思われる.
EcDOSは3′,5′-cyclic AMPの5′-AMPへの変換も行う.この反応は2価ヘム鉄の場合のみ進行し,3価ヘム鉄の場合反応は進行しないために,EcDOSはヘムレドックスセンサー(酸化還元センサー)ともいえる99, 109, 110).3′,5′-cyclic AMPに対する活性は0.15 min−1と非常に低いため,EcDOSの主な基質ではないと推定される.しかし,EcDOS遺伝子を破壊した大腸菌においてはその形態が変化するとともに,菌体内の3′,5′-cyclic AMPの濃度が26倍上昇することから,EcDOSは3′,5′-cyclic AMPが関わる代謝に何らかの寄与があると推定される111).
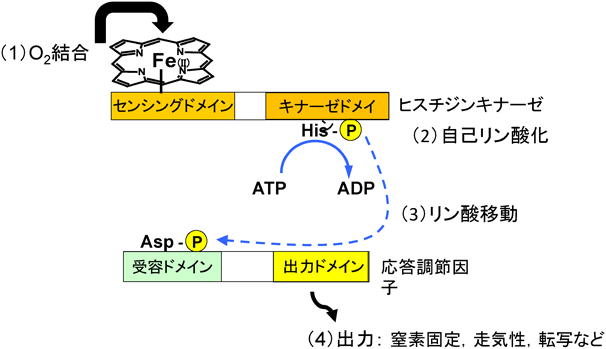
ヘム結合PAS構造ドメインをN末端に有する酸素センサーはこれ以外にFixLが重要である112–118).FixLは2成分シグナル伝達系(図8)を構成するヒスジンキナーゼである71).EcDOSと異なり,FixLの場合,センシングドメインの2価ヘム鉄に結合した酸素が解離することにより活性化し,まずヒスチジンが自己リン酸化する.ヒスチジン残基に付加されたリン酸基は,次いで応答調節因子FixJの受容ドメインにあるアスパラギン酸残基へ転移され,リン酸化されたFixJはニトロゲナーゼ遺伝子を活性化し,その発現を促進する.FixLの不活性型である2価ヘム鉄–酸素複合体[O2–Fe(II)–His]では,その酸素分子と遠位に位置するArg220が直接相互作用する点が,EcDOSの場合と類似している93)(図7B).AxPDEA1119)はPDE活性を有し,また,Aer2120)はメチル基受容走化性に関わるヘム結合PASドメインを有する酸素センサーである.
2)ヘム結合短縮グロビン構造ドメインを持つ酸素センサー
ヘモグロビンやミオグロビンなどにみられるタンパク質構造であるグロビン構造ドメインは,いくつかのαヘリックスより構成されている1, 2).細菌由来の酸素センサーの酸素センシングドメインにもヘム結合グロビンドメインが用いられる.しかし,酸素センサーのグロビンドメインは,動物のヘモグロビンなどの正常型グロビンドメインとは異なり,D-ヘリックスとE-ヘリックスの一部が欠損した短縮型グロビンドメイン構造をしている12, 91).短縮型グロビンドメインにヘム鉄が結合した細菌由来のヘム基盤酸素センサーはglobin-coupled oxygen sensor(GCS)とも呼ばれる.
大腸菌由来のYddV(EcDosC)はc-di-GMPを合成するGCSである(図6).酸素が2価ヘム鉄へ結合することによりc-di-GMPの合成は0.0 min−1から0.022 min−1へと上昇した121, 122).YddVの酸素の2価ヘム鉄への解離平衡定数Kdは14 μM程度で酸素の親和性は比較的高いことが判明した(表1).c-di-GMPを分解する同じ大腸菌由来のEcDOSのKdは340 μMであり酸素の親和性が低いことがわかっている.YddVのc-di-GMP合成活性回転数はEcDOSのc-di-GMP分解活性回転数に比べて低いが,YddVとEcDOSの自動酸化速度は同程度であり,大腸菌内の微小環境の一過的な酸素濃度に対応して,EcDOSとYddVが協同的に働き,c-di-GMPの濃度を調整していると考えられる(図6).実際に,c-di-GMPの原料であるGTPの水溶液に,この二つの酸素センサー酵素を混合すると,GTP→c-di-GMP→pGpG (l-di-GMP)→GMPのようにc-di-GMPの合成と分解の連続的な反応を観測することができた121).また,YddVを大腸菌に過剰発現させるとバイオフィルムの合成が確認されたことから,YddVの大腸菌における生理的役割の重要性が示唆された121).
これまでに結晶構造が決定されているGCSの一つであるHemATのヘム周辺構造とアミノ酸配列より,YddVでは2価ヘム鉄の遠位側にあるThr43が2価ヘム鉄–酸素複合体[O2–Fe(II)–His]の酸素分子と直接相互作用して,この酸素複合体を安定化していると推定された91, 121)(図7C).そのためにY43A, Y43L変異体では酸素の親和性が著しく低下し,酸素はこれらの変異体に結合しない.Y43W変異体では,酸素の解離速度定数,解離平衡定数,および自動酸化速度が著しく増加した(表1).また,共鳴ラマンスペクトルによって得られるYddVのO2–Fe(II)–His錯体のFe–O結合やO–O結合の伸縮振動数は,Y43F変異により著しく変化した121).また,ヘム鉄の遠位型に存在すると予想されるLeu65は水分子がヘム鉄へ接近することを妨げており,Leu65変異体では水分子が2価ヘム鉄へ接近し,その結果,自動酸化速度が著しく上昇し123),外部からの軸配位子の2価ヘム鉄への結合様式が変化した124, 125).Leu65変異体のヘム鉄周辺に存在する水分子は,ヘム酸素添加酵素(heme oxygenase)と同様に,分子状酸素からの酸素原子がヘム鉄のポルフィリンIXに添加されることを助けることが示唆された126, 127).
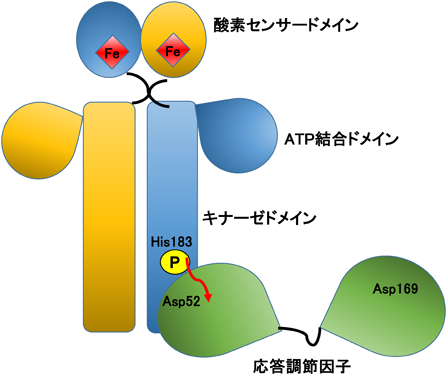
土壌細菌Anaeromyxobacter由来AfGcHKは2成分シグナル伝達系(図8)を構成するヘム基盤酸素センサー・キナーゼである71, 128, 129).同じ2成分シグナル伝達系のFixLでは酸素が2価ヘム鉄から解離することにより活性化するが,AfGcHKでは酸素が2価ヘム鉄へ結合することにより活性化する.酸素の結合により,まずHis183が自己リン酸化され,次いでそのリン酸が応答調節因子のAsp52およびAsp169へ転移する.近年開発されたPhos-tag SDS-PAGEゲル電気泳動法を活用したキナーゼ活性測定系を確立したことにより,放射性同位元素標識基質を使うこと無く,容易にリン酸化部位の同定やリン酸転移反応(キナーゼ反応)の反応速度論的解析が可能となった128, 129).本活性測定法を用いて,AfGcHKキナーゼ活性に関する反応速度論的パラメーターを詳細に検討した結果,酸素がAfGcHKのN末端のセンサードメインの2価ヘム鉄に結合することにより,C末端ドメインへのATPの親和性が高まり,その結果,自己リン酸化活性が上昇することが示唆された129).
GCSの一つであるHemATの構造とGCSのアミノ酸配列より,Tyr45が酸素結合錯体[O2–Fe(II)–His]の酸素分子と相互作用をして,この錯体の安定化に寄与すると推定された(図7C).Tyr45の変異体では,酸素の解離速度定数や解離平衡定数が野生型に比べて著しく増加し,Tyr45が酸素結合錯体の安定化に重要な役割を果たすことが確かめられた(表1).さらに,共鳴ラマンスペクトルより,Y45F変異体の酸素結合錯体O2–Fe(II)–HisのFe–O,およびO–O伸縮振動やそれらの重水素効果は野生型と著しく異なることが観測され,Tyr45が確かに酸素結合錯体の酸素原子と相互作用していることが強く示唆された.チロシンが2価ヘム鉄–酸素複合体の酸素と相互作用することは,別のGCSであるYddVと同様である(図7C).
タンパク質のX線結晶構造の決定が困難である場合,水素/重水素交換質量分析(hydrogen-deuterium exchange coupled to mass spectroscopy:HDX-MS)を用いると,水溶液中のタンパク質の構造やタンパク質–タンパク質相互作用が把握できる130).HDX-MSの結果より,AfGcHKのN末端のヘム結合短縮グロビン構造ドメインは硬く安定した構造をとってホモ二量体を形成し,ヘム鉄はグロビン構造の内部に埋め込まれていることが示された(図9).一方C末端は比較的柔軟な構造をしており単量体であると示唆された130).さらに,水溶液中で,応答調節因子の中で最初にリン酸化されるAsp52がAfGcHKの自己リン酸化部位であるHis183と近い位置に存在することが明らかになった(図9).
YddVやAfGcHKなどのGCSの酸素解離定数Kdは0.08~14 μMであり,EcDOSやFixLなどのPAS構造ドメイン酸素センサーのKd 12~500 μMと比べて低く,酸素親和性は高い91).このことより,これらのGCSは嫌気的条件下で薄い濃度の酸素を感知して機能すると推定される(表1).
GCSは他に,HemAT131–135),HemDGC136),HmAC-Lm137),PccGCS138),RsbR139),およびBpeGReg140)などがある.
3)ヘム結合GAF構造ドメインを持つ酸素センサー
GAF(cGMP-specific and -stimulated phosphodiesterase, adenylate cyclase, and E. coli formate hydrogen lyase transcriptional activator)構造ドメインはPAS構造と同様に主にβシートから構成されている141).ヘム鉄結合GAF構造を保持する酸素センサー,DosS(DevS)およびDosTも報告されている141–144).DosS, DosTともに,Fixと同様に2成分シグナル伝達系の一部になるヒスチジンキナーゼであり,酸素結合錯体が不活性型であり,酸素が解離することで活性化する(図8).
多くのヘム基盤酸素センサーのガス選択性は低い.すなわち,酸素センサーの2価ヘム鉄には酸素以外に,NO, COなども結合する場合が多い.しかし,ヘム基盤NOセンサーやヘム基盤COセンサーには,それぞれ,NOやCOが選択的に結合する場合が多い.これは,それぞれのガスの濃度が,細胞内で局所的,および過渡的に微妙に変化することと関係していると推定される.
ヘムセンサーとヘムレドックスセンサーの区別は明瞭ではない.ヘムセンサーで3価ヘム鉄と2価ヘム鉄との機能や配位構造の違いからヘムレドックスセンサーと示唆されている例もある37, 38, 54, 68).しかし,これらのヘム鉄の還元過程は研究されておらず,好気的条件下では2価ヘム鉄–O2結合体が生じるし,これは容易に自動酸化されて3価ヘム鉄へ変換する可能性も高い.
ヘムセンサーかつヘム基盤COセンサーには解決すべき問題が残っている.たとえば,NPAS240),Rev-erbβ52, 68),K+チャネル58–60, 176, 177),E7553),PGRMC175)などでは,それらの機能が3価ヘム鉄の結合・解離により制御され,かつCOによって制御される「ヘムセンサーかつヘム基盤COセンサー」と報告されている.しかし,COはタンパク質やアミノ酸と反応せず,金属のみと反応して金属カルボニルを合成する161, 178).特にヘム鉄ではCOは2価ヘム鉄にのみ結合し,3価ヘム鉄とは反応しない.一般に,ヘムタンパク質,特にヘムセンサーやヘム転送タンパク質では,3価ヘム鉄の配位構造(軸配位子としてのシステインまたはチロシン)やヘム周辺構造は2価ヘム鉄へ還元された時点で著しく変化するが,それについて十分に検討されていない.また,これらのCOセンサーの多くでは,3価ヘム鉄を2価ヘム鉄へ還元する還元物質や還元酵素が知られていない.一方,COが生理作用に影響を及ぼすとの報告にも,COと直接相互作用する標的がヘム鉄か他の金属イオンかについては明らかではない179, 180).
ヘムセンサーやヘム基盤ガスセンサーについて,細胞・組織レベルの実験系において観測された現象が,精製標品を対象にして生化学的および物理化学的実験から得られたデータに基づいて積み上げられた理論では矛盾なく説明することが難しい場合もあり,さらに詳細な分子機構の解明に興味が待たれる.今後も,特に疾病に関係する,多くの興味ある重要な結果が報告されることが期待される.
引用文献References
1) Poulos, T.L. (2014) Chem. Rev., 114, 3919–3962.
2) Voet, D. & Voet, J.G.(2011) In Biochemistry 4th ed., pp. 323–358, John Wiley & Sons, New York.
3) Reddi, A.R. & Hamza, I. (2016) Acc. Chem. Res., 49, 1104–1110.
4) Roumenina, L.T., Rayes, J., Lacroix-Desmazes, S., & Demitrov, J.D. (2016) Trends Mol. Med., 22, 200–213.
5) Haldar, S.M. & Stamler, J.S. (2013) J. Clin. Invest., 123, 101–110.
6) Yukl, E.T., de Vries, S., & Moënne-Loccoz, P. (2009) J. Am. Chem. Soc., 131, 7234–7235.
7) Corti, P., Xue, J., Tejero, J., Wajih, N., Sun, M., Stolz, D.B., Tsang, M., Kim-Shapiro, D.B., & Gladwin, M.T. (2016) Proc. Natl. Acad. Sci. USA, 113, 8538–8543.
8) Hannibal, L., Tomasina, F., Capdevila, D.A., Demicheli, V., Tortóra, V., Alvarez-Raggi, D., Jemmerson, R., Murgida, D.H., & Radi, R. (2016) Biochemistry, 55, 407–428.
9) Shimizu, T. (2012) J. Inorg. Biochem., 108, 171–177.
10) Igarashi, J., Kitanishi, K., Martinkova, M., Murase, M., Iizuka, A., & Shimizu, T. (2008) Acta Chim. Slov., 55, 67–74.
11) Igarashi, J., Kitanishi, K., & Shimizu, T. (2011) in Handbook of Porphyrin Science (Kadish, K. M., Smith, K. M., & Guilard, R. eds), Vol. 15, Chapter 73, pp. 399–460, World Scientific Publishing, Hackensack, NJ.
12) Shimizu, T., Huang, D., Yan, F., Stranava, M., Bartosoa, M., Fojtíková, V., & Martínková, M. (2015) Chem. Rev., 115, 6491–6533.
13) Hanna, D.A., Harvey, R.M., Martinez-Guzman, O., Yuan, X., Chandrasekharan, B., Raju, G., Outten, J.W., Hamza, I., & Reddi, A.R. (2016) Proc. Natl. Acad. Sci. USA, 113, 7539–7544.
14) Holcik, M. & Sonenberg, N. (2005) Nat. Rev. Mol. Cell Biol., 6, 318–327.
15) Chen, J.J. (2000) in Translational Control of Gene Expression (Soneberg, N., Hershey, J.W.B., & Mathews, M.B. eds.), pp. 529–546, Cold Spring Harbor Laboratory, Cold Spring Harbor, NY.
16) Zhang, L. & Guarente, L. (1995) EMBO J., 14, 313–320.
17) Igarashi, J., Sato, A., Kitagawa, T., Yoshimura, T., Yamauchi, S., Sagami, I., & Shimizu, T. (2004) J. Biol. Chem., 279, 15752–15762.
18) Miksanova, M., Igarashi, J., Minami, M., Sagami, I., Yamauchi, S., Kukokawa, H., & Shimizu, T. (2006) Biochemistry, 45, 9894–9905.
19) Igarashi, J., Murase, M., Iizuka, A., Pichierri, F., Martinkova, M., & Shimizu, T. (2008) J. Biol. Chem., 283, 18782–18791.
20) Carter, E.L., Gupta, N., & Ragsdale, S.W. (2016) J. Biol. Chem., 291, 2196–2222.
21) Schneider, S., Sharp, K.H., Barker, P.D., & Paoli, M. (2006) J. Biol. Chem., 281, 32606–32610.
22) Watanabe-Matsui, M., Matsumoto, T., Matsui, T., Ikeda-Saito, M., Muto, A., Murayama, K., & Igarashi, K. (2015) Arch. Biochem. Biophys., 565, 25–31.
23) Burton, M.J., Kapetanaki, S.M., Chernova, T., Jamieson, A.G., Dorlet, P., Santolini, J., Moody, P.C.E., Mitcheson, J.S., Davies, N.W., Schmid, R., Raven, E.L., & Storey, N.M. (2016) Proc. Natl. Acad. Sci. USA, 113, 3785–3790.
24) Igarashi, J., Sasaki, T., Kobayashi, N., Yoshioka, S., Matsushita, M., & Shimizu, T. (2011) FEBS J., 278, 918–928.
25) Martinkova, M., Igarashi, J., & Shimizu, T. (2007) FEBS Lett., 581, 4109–4114.
26) Faller, M., Matsunaga, M., Yin, S., Loo, J.A., & Guo, F. (2007) Nat. Struct. Mol. Biol., 14, 23–29.
27) Quick-Cleveland, J., Jacob, J.P., Weitz, S.H., Shoffner, G., Senturia, R., & Guo, F. (2014) Cell Reports, 7, 1994–2005.
28) Girvan, H.M., Bradley, J.M., Cheesman, M.R., Kincaid, J.R., Liu, Y., Czarnecki, K., Fisher, K., Ley, D., Rigby, S.E.J., & Munro, A.W. (2016) Biochemistry, 55, 5073–5083.
29) Kubota, Y., Nomura, K., Katoh, Y., Yamashita, R., Kaneko, K., & Furuyama, K. (2016) J. Biol. Chem., 291, 20516–20529.
30) Igarashi, K. & Watanabe-Matsui, M. (2014) Tohoku J. Exp. Med., 232, 229–253.
31) Ogawa, K., Sun, J., Taketani, S., Nakajima, O., Nishitani, C., Sassa, S., Hayashi, N., Yamamoto, M., Shibahara, S., Fujita, H., & Igarashi, K. (2001) EMBO J., 20, 2835–2843.
32) Sun, J., Hoshino, H., Takaku, K., Nakajima, O., Muto, A., Suzuki, H., Tashiro, S., Takahashi, S., Shibahara, S., Alam, J., Taketo, M.M., Yamamoto, M., & Igarashi, K. (2002) EMBO J., 21, 5216–5224.
33) Hira, S., Tomita, T., Matsui, T., Igarashi, K., & Ikeda-Saito, M. (2007) IUBMB Life, 59, 542–551.
34) Watanabe-Matsui, M., Muto, A., Matsui, T., Itoh-Nakadai, A., Nakajima, O., Murayama, K., Yamamoto, M., Ikeda-Saito, M., & Igarashi, K. (2011) Blood, 117, 5438–5448.
35) Shen, J., Sheng, X., Chang, Z., Wu, Q., Wang, S., Xuan, Z., Li, D., Wu, Y., Shang, Y., Kong, X., Yu, L., Li, L., Ruan, K., Hu, H., Huang, Y., Hui, L., Xie, D., Wang, F., & Hu, R. (2014) Cell Reports, 7, 180–193.
36) Mourer, T., Jacques, J.-F., Brault, A., Bisaillon, M., & Labbé, S. (2015) J. Biol. Chem., 290, 10176–10190.
37) Yin, L., Dragnea, V., & Bauer, C.E. (2012) J. Biol. Chem., 287, 13850–13858.
38) Tang, K., Knipp, M., Liu, B.-B., Cox, N., Stabel, R., He, Q., Zhou, M., Scheer, H., Zhao, K.-H., & Gärtner, W. (2015) J. Biol. Chem., 290, 19067–19080.
39) Laakso, H.A., Marolda, C.L., Pinter, T.B., Stillman, M.J., & Heinrichs, D.E. (2016) J. Biol. Chem., 291, 29–40.
40) Dioum, E.M., Rutter, J., Tuckerman, J.R., Gonzalez, G., Gilles-Gonzalez, M.A., & McKnight, S.L. (2002) Science, 298, 2385–2387.
41) Kitanishi, K., Hayasaka, K., Harada, M., Nagata, R., Igarashi, J., & Shimizu, T. (2011) in Circadian Rhythms: Biology, Cognition and Disorders (Golovkin, L., & Maliszkewicz, A. eds.), pp. 225–245, Nova Sci. Publ., NY.
42) Uchida, T., Sato, E., Sato, A., Sagami, I., Shimizu, T., & Kitagawa, T. (2005) J. Biol. Chem., 280, 21358–21368.
43) Mukaiyama, Y., Uchida, T., Sato, E., Sasaki, A., Sato, Y., Igarashi, J., Kurokawa, H., Sagami, I., Kitagawa, T., & Shimizu, T. (2006) FEBS J., 273, 2528–2539.
44) Ishida, M., Ueha, T., & Sagami, I. (2008) Biochem. Biophys. Res. Commun., 368, 292–297.
45) Yoshii, K., Tajima, F., Ishijima, S., & Sagami, I. (2015) Biochemistry, 54, 250–259.
46) Yoshii, K., Ishijima, S., & Sagami, I. (2013) Biochem. Biophys. Res. Commun., 437, 386–391.
47) Nagata, R., Harada, M., Kitanishi, K., Igarashi, J., Uchida, T., Ishimori, K., & Shimizu, T. (2011) in Circadian Rhythms: Biology, Cognition and Disorders (Golovkin, L., Maliszkewicz, A. eds.), pp. 133–160, Nova Sci. Publ., NY.
48) Kaasik, K. & Lee, C.C. (2004) Nature, 430, 467–471.
49) Kitanishi, K., Igarashi, J., Hayasaka, K., Hikage, N., Saiful, I., Yamauchi, S., Uchida, T., Ishimori, K., & Shimizu, T. (2008) Biochemistry, 47, 6157–6168.
50) Hayasaka, K., Kitanishi, K., Igarashi, J., & Shimizu, T. (2011) Biochim. Biophys. Acta, 1814, 326–333.
51) Yang, J., Kim, K.D., Lucas, A., Drahos, K.E., Santos, C.S., Mury, S.P., Capelluto, D.G.S., & Finkielstein, C.V. (2008) Mol. Cell. Biol., 28, 4697–4711.
52) Gupta, N. & Ragsdale, S.W. (2011) J. Biol. Chem., 286, 4392–4403.
53) Marvin, K.A., Reinking, J.L., Lee, A.J., Pardee, K., Krause, H.M., & Burstyn, J.N. (2009) Biochemistry, 48, 7056–7071.
54) Reinking, J., Lam, M.M.S., Pardee, K., Sampson, H.M., Liu, S., Yang, P., Williams, S., White, W., Lajoie, G., Edwards, A., & Krause, H.M. (2005) Cell, 122, 195–207.
55) de Rosny, E., de Groot, A., Jullian-Binard, C., Gaillard, J., Borel, F., Pebay-Peyroula, E., Fontecilla-Camps, J.C., & Jouve, H. (2006) Biochemistry, 45, 9727–9734.
56) Sahoo, N., Goradia, N., Ohlenshläger, O., Schönherr, R., Friedrich, M., Plass, W., Kappl, R., Hoshi, T., & Heinemann, S.H. (2013) Proc. Natl. Acad. Sci. USA, 110, E4036–E4044.
57) Tang, X.D., Xu, R., Reynolds, M.F., Carcia, M.L., Heinemann, S.H., & Hoshi, T. (2003) Nature, 425, 531–535.
58) Hou, S., Reynolds, M.F., Horrigan, F.T., Heinemann, S.H., & Hoshi, T. (2006) Acc. Chem. Res., 39, 918–924.
59) Yi, L., Morgan, J.T., & Ragsdale, S.W. (2010) J. Biol. Chem., 285, 20117–20127.
60) Williams, S.E.J., Wootton, P., Mason, H.S., Bould, J., Iles, D.E., Riccardi, D., Peers, C., & Kemp, P.J. (2004) Science, 306, 2093–2097.
61) Ishikawa, H., Kato, M., Hori, H., Ishimori, K., Kirisako, T., Tokunaga, F., & Iwai, K. (2005) Mol. Cell, 19, 171–181.
62) Kitatsuji, C., Izumi, K., Nambu, S., Kurogochi, M., Uchida, T., Nishimura, S., Iwai, K., O’Brian, M.R., Ikeda-Saito, M., & Ishimori, K. (2016) Sci. Rep., 6, 18703.
63) Ishikawa, H., Nakagaki, M., Bamba, A., Uchida, T., Hori, H., O’Brian, M.R., Iwai, K., & Ishimori, K. (2011) Biochemistry, 50, 1016–1022.
64) Kobayashi, K., Nakagaki, M., Ishikawa, H., Iwai, K., O’Brian, M.R., & Ishimori, K. (2016) Biochemistry, 55, 4047–4054.
65) Uchida, T., Sekine, Y., Matsui, T., Ikeda-Saito, M., & Ishimori, K. (2012) Chem. Commun. (Camb.), 48, 6741–6743.
66) Yin, L., Wu, N., Curtin, J.C., Qatanami, M., Szwergold, N.R., Reid, R.A., Waitt, G.M., Parks, D.J., Pearce, K.H., Wisely, G.B., & Lazar, M.A. (2007) Science, 318, 1786–1789.
67) Raghuram, S., Stayrook, K.R., Huang, P., Rogers, P.M., Kosie, A.K., McClure, D.B., Burris, L.L., Khorasanizadeh, S., Burris, T.P., & Rastinejad, F. (2007) Nat. Struct. Mol. Biol., 14, 1207–1213.
68) Pardee, K.I., Xu, X., Reiking, J., Schuez, A., Dong, A., Liu, S., Zhang, R., Tiefenbach, J., Lajoie, G., Plotnikov, A.N., Botchkarev, A., Krause, H.M., & Edwards, A. (2009) PLoS Biol., 7, e1000043.
69) Sawai, H., Yamanaka, M., Sugimoto, H., Shiro, Y., & Aono, S. (2012) J. Biol. Chem., 287, 30755–30768.
70) Wakasugi, K. (2007) Biochemistry, 46, 11291–11298.
71) Casino, P., Rubio, V., & Marina, A. (2010) Curr. Opin. Struct. Biol., 20, 763–771.
72) Ito, Y., Nakagawa, S., Komagata, A., Ikeda-Saito, M., Shiro, Y., & Nakamura, H. (2009) FEBS Lett., 583, 2244–2248.
73) Bagai, I., Sarangi, R., Fleischhacker, A.S., Sharm, A., Hoffman, B.M., Zuiderweg, E.R.P., & Ragsdale, S.W. (2015) Biochemistry, 54, 2693–2708.
74) Fleischhacker, A.S., Sharma, A., Choi, M., Spencer, A.M., Bagai, I., Hoffman, B.M., & Ragsdale, S.W. (2015) Biochemistry, 54, 2709–2718.
75) Kabe, Y., Nakane, T., Koike, I., Yamamoto, T., Sugiura, Y., Harada, E., Sugase, K., Shimamura, T., Ohmura, M., Muraoka, K., Yamamoto, A., Uchida, T., Iwata, S., Yamaguchi, Y., Krahukhina, E., Noda, M., Handa, H., Ishimori, K., Uchiyama, S., Kobayashi, T., & Suematsu, M. (2016) Nat. Commun., 7, 11030.
76) Kanadani, M., Sato, T., Hino, T., Nagano, S., & Ozaki, S. (2015) J. Inorg. Biochem., 151, 26–33.
77) Watanabe, M., Tanaka, Y., Suenaga, A., Kuroda, M., Yao, M., Watanabe, N., Aisaka, F., Ohta, T., Tanaka, I., & Tsumoto, K. (2008) J. Biol. Chem., 283, 28649–28659.
78) Sekine, Y., Tanzawa, T., Tanaka, Y., Ishimori, K., & Uchida, T. (2016) Biochemistry, 55, 884–893.
79) Moriwaki, Y., Terada, T., Caaveiro, J.M.M., Takaoka, Y., Hamachi, I., Tsumoto, K., & Shimizu, K. (2013) Biochemistry, 52, 8866–8877.
80) Ozaki, S., Sato, T., Sekine, Y., Migita, C.T., Uchida, T., & Ishimori, K. (2014) J. Inorg. Biochem., 138, 31–38.
81) Smith, A.D., Modi, A.R., Sun, S., Dawson, J.H., & Wilks, A. (2015) Biochemistry, 54, 2601–2612.
82) Draganova, E.B., Akbas, N., Adrian, S.A., Lukat-Rodgers, G.S., Collins, D.P., Dawson, J.H., Allen, C.E., Schmitt, M.P., Rodgers, K.R., & Dixon, D.W. (2015) Biochemistry, 54, 6598–6609.
83) Burkhard, K.A. & Wilks, A. (2007) J. Biol. Chem., 282, 15126–15136.
84) Cobessi, D., Meksem, A., & Brillet, K. (2009) Proteins, 78, 286–294.
85) Piel, R.B. III, Shiferaw, M.T., Vashisht, A.V., Marcero, J.R., Praissman, J.L., Phillips, J.D., Wohlschlegel, J.A., & Medlock, A.E. (2016) Biochemistry, 55, 5204–5217.
86) Kaluka, D., Batabyal, D., Chiang, B.-Y., Poulos, T.L., & Yeh, S.-R. (2015) Biochemistry, 54, 1638–1647.
87) Kobayashi, K., Yoshioka, S., Kato, Y., Asano, Y., & Aono, S. (2005) J. Biol. Chem., 280, 5486–5490.
88) Sawai, H., Sugimoto, H., Kato, Y., Asano, Y., Shiro, T., & Aono, S. (2009) J. Biol. Chem., 284, 32089–32096.
89) Beltrán, J., Kloss, B., Hosler, J.P., Geng, J., Liu, A., Modi, A., Dawson, J.H., Sono, M., Shumskaya, M., Ampomah-Dwamena, C., Love, J.D., & Wurtzel, E.T. (2015) Nat. Chem. Biol., 11, 598–605.
90) Uchida, T. & Kitagawa, T. (2005) Acc. Chem. Res., 38, 662–670.
91) Martínková, M., Kitanishi, K., & Shimizu, T. (2013) J. Biol. Chem., 288, 27702–27711.
92) Henry, J.T. & Crosson, S. (2011) Annu. Rev. Microbiol., 65, 261–286.
93) Hao, B., Isaza, C., Arndt, J., Soltis, M., & Chan, M.K. (2002) Biochemistry, 41, 12952–12958.
94) Kurokawa, H., Lee, D.S., Watanabe, M., Sagami, I., Mikami, B., Raman, C.S., & Shimizu, T. (2004) J. Biol. Chem., 279, 20186–20193.
95) Park, H., Suquet, C., Satterlee, J.D., & Kang, C. (2004) Biochemistry, 43, 2738–2746.
96) Schirmer, T. & Jenal, U. (2009) Nat. Rev. Microbiol., 7, 724–735.
97) Delgano-Nixon, V.M., Gonzalez, G., & Gilles-Gonzalez, M.A. (2000) Biochemistry, 39, 2685–2691.
98) Shimizu, T. (2013) Biosensor, 3, 211–237.
99) Sasakura, Y., Yoshimura-Suzuki, T., Kurokawa, H., & Shimizu, T. (2006) Acc. Chem. Res., 39, 37–43.
100) Tanaka, A., Takahashi, H., & Shimizu, T. (2007) J. Biol. Chem., 282, 21301–21307.
101) Tanaka, A. & Shimizu, T. (2008) Biochemistry, 47, 13438–13446.
102) Ishitsuka, Y., Araki, Y., Tanaka, A., Igarashi, J., Ito, O., & Shimizu, T. (2008) Biochemistry, 47, 8874–8884.
103) Taguchi, S., Matsui, T., Igarashi, J., Sasakura, Y., Araki, Y., Ito, O., Sugiyama, S., Sagami, I., & Shimizu, T. (2004) J. Biol. Chem., 279, 3340–3347.
104) Yamashita, T., Bouzhir-Sima, L., Lambry, J.C., Liebl, U., & Vos, M.H. (2008) J. Biol. Chem., 283, 2344–2352.
105) Hirata, S., Matsui, T., Sasakura, Y., Sugiyama, S., Yoshimura, T., Sagami, I., & Shimizu, T. (2003) Eur. J. Biochem., 270, 4771–4779.
106) Sato, A., Sasakura, Y., Sugiyama, S., Sagami, I., Shimizu, T., Mizutani, Y., & Kitagawa, T. (2002) J. Biol. Chem., 277, 32650–32658.
107) El-Mashtoly, S.F., Nakashima, S., Tanaka, A., Shimizu, T., & Kitagawa, T. (2008) J. Biol. Chem., 283, 19000–19010.
108) El-Mashtoly, S.F., Takahashi, H., Shimizu, T., & Kitagawa, T. (2007) J. Am. Chem. Soc., 129, 3556–3563.
109) Sasakura, Y., Hirata, S., Sugiyama, S., Suzuki, S., Taguchi, S., Watanabe, M., Matsui, T., Sagami, I., & Shimizu, T. (2002) J. Biol. Chem., 277, 23821–23827.
110) Yoshimura, T., Sagami, I., Sasakura, Y., & Shimizu, T. (2003) J. Biol. Chem., 278, 53105–53111.
111) Yoshimura-Suzuki, T., Sagami, I., Yokota, N., Kurokawa, H., & Shimizu, T. (2005) J. Bacteriol., 187, 6678–6682.
112) Gilles-Gonzalez, M.A., Ditta, G.S., & Helinski, D.R. (1991) Nature, 350, 170–172.
113) Gilles-Gonzalez, M.A., Gonzalez, G., Perutz, M.F., Kiger, L., Marden, M.C., & Poyart, C. (1994) Biochemistry, 33, 8067–8073.
114) Miyatake, H., Mukai, M., Adachi, S., Nakamura, H., Tamura, K., Iizuka, T., Shiro, Y., Strange, R.W., & Hasnain, S.S. (1999) J. Biol. Chem., 274, 23176–23184.
115) Miyatake, H., Mukai, M., Park, S.-Y., Adachi, S., Tamura, K., Nakamura, H., Nakamura, K., Tsuchiya, T., Iizuka, T., & Shiro, Y. (2000) J. Mol. Biol., 301, 415–431.
116) Nakamura, H., Kumita, H., Imai, K., Iizuka, T., & Shiro, Y. (2004) Proc. Natl. Acad. Sci. USA, 101, 2742–2746.
117) Tanaka, A., Nakamura, H., Shiro, Y., & Fujii, H. (2006) Biochemistry, 45, 2515–2523.
118) Yamawaki, T., Ishikawa, H., Mizuno, M., Nakamura, H., Shiro, Y., & Mizutani, Y. (2016) Biochemistry, 55, 4027–4035.
119) Chang, A.L., Tuckerman, J.R., Gonzalez, G., Mayer, R., Weinhouse, H., Volman, G., Amikam, D., Benziman, M., & Gilles-Gonzalez, M.A. (2001) Biochemistry, 40, 3420–3426.
120) Sawai, H., Sugimoto, H., Shiro, Y., Ishikawa, H., Mizutani, Y., & Aono, S. (2012) Chem. Commun. (Camb.), 48, 6523–6525.
121) Kitanishi, K., Kobayashi, K., Kawamura, Y., Ishigami, I., Ogura, T., Nakajima, K., Igarashi, J., Tanaka, A., & Shimizu, T. (2010) Biochemistry, 49, 10381–10393.
122) Tuckerman, J.R., Gonzalez, G., Sousa, E.H.S., Wan, X., Saito, J.A., Alam, M., & Gilles-Gonzalez, M.A. (2009) Biochemistry, 48, 9764–9774.
123) Nakajima, K., Kitanishi, K., Kobayashi, K., Kobayashi, N., Igarashi, J., & Shimizu, T. (2012) J. Inorg. Biochem., 108, 163–170.
124) Lambry, J.C., Stranava, M., Lobato, L., Martinkova, M., Shimizu, T., Lieble, U., & Vos, M.H. (2016) J. Phys. Chem. Lett., 7, 69–74.
125) Pavlou, A., Martínkova, M., Shimizu, T., Kitanishi, K., Stranava, M., Loullis, A., & Pinakoulaki, E. (2015) Phys. Chem. Chem. Phys., 17, 17007–17015.
126) Stranava, M., Martínková, M., Stiborová, M., Man, P., Kitanishi, K., Muchová, L., Vítek, L., Martínek, V., & Shimizu, T. (2014) J. Inorg. Biochem., 140, 29–38.
127) Matsui, T., Iwasaki, M., Sugiyama, R., Unno, M., & Ikeda-Saito, M. (2010) Inorg. Chem., 49, 3602–3609.
128) Kitanishi, K., Kobayashi, K., Uchida, T., Ishimori, K., Igarashi, J., & Shimizu, T. (2011) J. Biol. Chem., 286, 35522–35534.
129) Fojtikova, V., Stranava, M., Vos, M.H., Liebl, U., Hranicek, J., Kitanishi, K., Shimizu, T., & Martinkova, M. (2015) Biochemistry, 54, 5017–5029.
130) Stranava, M., Martínek, V., Man, P., Fojtikova, V., Kavan, D., Vaněk, O., Shimzu, T., & Martinkova, M. (2016) Proteins, 84, 1375–1389.
131) Hou, S., Larsen, R.W., Boudko, D., Riley, C.W., Karatan, E., Zimmer, M., Ordal, G.W., & Alam, M. (2000) Nature, 403, 540–544.
132) Hou, S., Freitas, T., Larsen, R.W., Piatibratov, M., Sivozhelezov, V., Yamamoto, A., Meleshkevitch, E.A., Zimmer, M., Ordal, G.W., & Alam, M. (2001) Proc. Natl. Acad. Sci. USA, 98, 9353–9358.
133) El-Mashtoly, S.F., Kubo, M., Gu, Y., Sawai, H., Nakajima, S., Ogura, T., Aono, S., & Kitagawa, T. (2012) J. Biol. Chem., 287, 19973–19984.
134) Yoshimura, H., Yoshioka, S., Kobayashi, K., Ohta, T., Uchida, T., Kubo, M., Kitagawa, T., & Aono, S. (2006) Biochemistry, 45, 8301–8307.
135) Aono, S., Kato, T., Matsuki, M., Nakajima, H., Ohta, T., Uchida, T., & Kitagawa, T. (2002) J. Biol. Chem., 277, 13528–13538.
136) Sawai, H., Yoshioka, S., Uchida, T., Hyodo, M., Hayakawa, Y., Ishimori, K., & Aono, S. (2010) Biochim. Biophys. Acta, 1804, 166–172.
137) Santara, S.S., Roy, J., Mukherjee, S., Rose, M., Saha, R., & Adak, S. (2013) Proc. Natl. Acad. Sci. USA, 110, 16790–16795.
138) Burns, J.L., Deer, D.D., & Weinert, E.E. (2014) Mol. Biosyst., 10, 2823–2826.
139) Jia, X., Wang, J., Rivera, S., Duong, D., & Weinert, E.E. (2016) Nat. Commun., 7, 12381.
140) Burns, J.L., Rivera, S., Deer, D.D., Joynt, S.C., Dvorak, D., & Weinert, E.E. (2016) Biochemistry, 55, 6642–6651.
141) Podust, L.M., Ioanoviciu, A., & Ortiz de Montellano, P.R. (2008) Biochemistry, 47, 12523–12531.
142) Kumar, A., Toledo, J.C., Patel, R.P., Lancaster, J.R. Jr., & Steyn, A.J.C. (2007) Proc. Natl. Acad. Sci. USA, 104, 11568–11573.
143) Sousa, E.H.S., Tuckerman, J.R., Gonzalez, G., & Gilles-Gonzalez, M.A. (2007) Protein Sci., 16, 1708–1719.
144) Basudhar, D., Madrona, Y., Yukl, E.T., Sivaramakrishnan, S., Nishida, C.R., Moënne-Loccoz, P., & Ortiz de Montellano, P.R. (2016) J. Biol. Chem., 291, 16100–16111.
145) Cary, S.P.L., Winger, J.A., Derbyshire, E.R., & Marletta, M.A. (2006) Trends Biochem. Sci., 31, 231–239.
146) Sagami, I., Daff, S., & Shimizu, T. (2001) J. Biol. Chem., 276, 30036–30042.
147) Sato, Y., Sagami, I., & Shimizu, T. (2004) J. Biol. Chem., 279, 8827–8836.
148) Derbyshire, E.R. & Marletta, M.A. (2012) Annu. Rev. Biochem., 81, 533–559.
149) Yazawa, S., Tsuchiya, H., Hori, H., & Makino, R. (2006) J. Biol. Chem., 281, 21763–21770.
150) Makino, R., Park, S.-Y., Obayashi, E., Iizuka, T., Hori, H., & Shiro, Y. (2011) J. Biol. Chem., 286, 15678–15687.
151) Makino, R., Yazawa, S., Hori, H., & Shiro, Y. (2012) Biochemistry, 51, 9277–9289.
152) Martin, E., Berka, V., Bogatenkova, E., Murad, F., & Tsai, A.-L. (2006) J. Biol. Chem., 281, 27836–27845.
153) Stone, J.R. & Marketta, M.A. (1994) Biochemistry, 33, 5636–5640.
154) Plate, L. & Marletta, M.A. (2013) Trends Biochem. Sci., 38, 566–575.
155) Nisbett, L.-M. & Boon, E.M. (2016) Biochemistry, 55, 4873–4884.
156) Takeda, T., Mu, A., Tai, T.T., Kajima, S., & Taketani, S. (2015) Sci. Rep., 5, 10488.
157) Liem, P.H., Mu, A., Kikuta, S., Ohta, K., Kitajima, S., & Taketani, S. (2015) Biol. Chem., 396, 1265–1268.
158) Matsui, T., Nambu, S., Goulding, C.W., Takahashi, S., Fujii, H., & Ikeda-Saito, M. (2016) Proc. Natl. Acad. Sci. USA, 113, 3779–3784.
159) Sugishima, H., Sato, H., Higashimoto, Y., Harada, J., Wada, K., Fukuyama, K., & Noguchi, M. (2014) Proc. Natl. Acad. Sci. USA, 111, 2524–2529.
160) Sugishima, M., Moffat, K., & Noguchi, M. (2012) Biochemistry, 51, 8554–8562.
161) Bürstel, I., Siebert, E., Frielingsdorf, S., Zebger, I., Friedrich, B., & Lenz, O. (2016) Proc. Natl. Acad. Sci. USA, 113, 14722–14726.
162) Shelver, D., Kerby, R.L., He, Y., & Roberts, G.P. (1997) Proc. Natl. Acad. Sci. USA, 94, 11216–11220.
163) Aono, S. (2003) Acc. Chem. Res., 36, 825–831.
164) Roberts, G.P., Kerby, R.L., Youn, H., & Conrad, M. (2005) J. Inorg. Biochem., 99, 280–292.
165) Lanzilotta, W.N., Schuller, D.J., Thorsteinsson, M.V., Kerby, R.L., Roberts, G.P., & Poulos, T.L. (2000) Nat. Struct. Biol., 7, 876–880.
166) Yamashita, T., Hoashi, Y., Watanabe, K., Tomisugi, Y., Ishikawa, Y., & Uno, T. (2004) J. Biol. Chem., 279, 21394–21400.
167) Yamashita, T., Hoashi, Y., Tomisugi, Y., Ishikawa, Y., & Uno, T. (2004) J. Biol. Chem., 279, 47320–47325.
168) Kubo, M., Inagaki, S., Yoshioka, S., Uchida, T., Mizutani, Y., Aono, S., & Kitagawa, T. (2006) J. Biol. Chem., 281, 11271–11278.
169) Komori, H., Inagaki, S., Yoshioka, S., Aono, S., & Higuchi, Y. (2007) J. Mol. Biol., 367, 864–871.
170) Inagaki, S., Masuda, C., Akaishi, T., Nakajima, H., Yoshioka, S., Ohta, T., Pal, B., Kitagawa, T., & Aono, S. (2005) J. Biol. Chem., 280, 3269–3274.
171) Singh, S. & Banerjee, R. (2011) Biochim. Biophys. Acta, 1814, 1518–1527.
172) Kabil, O. & Banerjee, R. (2010) J. Biol. Chem., 285, 21903–21907.
173) Yadav, P.K., Xie, P., & Banerjee, R. (2012) J. Biol. Chem., 287, 37611–37620.
174) Vicente, J.B., Colaço, H.G., Mendes, M.I.S., Sarti, P., Leandro, P., & Giuffrè, A. (2014) J. Biol. Chem., 289, 8579–8587.
175) Kabil, O., Weeks, C.L., Carballal, S., Gherasim, C., Alvarez, B., Spiro, T.G., & Banerjee, R. (2011) Biochemistry, 50, 8261–8263.
176) Hou, S., Xu, R., Haineman, S.H., & Hoshi, T. (2008) Proc. Natl. Acad. Sci. USA, 105, 4039–4043.
177) Al-Owais, M.M.A., Scragg, J.L., Dallas, M.L., Boycott, H.E., Warburton, P., Chakrabarty, A., Boyle, J.P., & Peers, C. (2012) J. Biol. Chem., 287, 24754–24764.
178) Spatzal, T., Perez, K.A., Einsle, O., Howard, J.B., & Rees, D.C. (2014) Science, 345, 1620–1623.
179) Elies, J., Dallas, M.L., Boyle, J.P., Scragg, J.L., Duke, A., Steel, D.S., & Peers, C. (2014) J. Biol. Chem., 289, 16421–16429.
180) Klemz, R., Reischl, S., Wallach, T., Witte, N., Jürchotte, K., Klemz, S., Lang, V., Lorenzen, S., Knauer, M., Heidenreich, S., Xu, M., Ripperger, J.A., Schupp, M., Stanewsky, R., & Kramer, A. (2016) Nat. Struct. Mol. Biol., 24, 15–22.
181) Honorio-Felício, N., Carepo, M.S.P., Paulo, T.F., Gonzaga, L., Lopes, L.G.F., Sousa, E.H.S., Diógenes, I.C.N., & Bernhardt, P.V. (2016) J. Inorg. Biochem., 164, 34–41.