嗅覚系は物体から発せられる匂い分子を受容し,その情報を鼻から脳へと伝え,匂いのイメージを脳内に表現する神経システムである.また鋤鼻系は同種の他個体から発せられるフェロモン分子を受容し,内分泌系の変化や特有の行動を発現させる神経システムである.これらの感覚は総称して「化学感覚」と呼ばれ,無脊椎動物から哺乳類に至るまで多くの生物種で重要な役割を果たしている.化学感覚の入力情報をもとにして,物体の認知,快・不快という感情の誘起,記憶の形成・想起,内分泌系の変化などが起こり,個体の生存,生体の恒常性維持や種の保存のために必要な行動出力が発現・調節される.とりわけ,好きな匂いへの誘引行動,嫌いな匂いからの逃避行動,性フェロモンを介した性行動は,多くの生物に共通する三つの根源的な嗅覚行動である1).
キンギョなどの魚類において,脂質メディエーターであるプロスタグランジンF2α(PGF2α)は,メスの体内で排卵・産卵を調節するホルモンとして働くだけでなく,メスから水中に放出されたPGF2αがオスの性行動を誘起する性フェロモンとしても機能することが1980年代に報告されていた2).しかしながら,その神経回路メカニズムについてはこれまでまったくわかっていなかった.我々はゼブラフィッシュを用いて,性フェロモンPGF2αを認識する嗅覚受容体の同定および求愛行動促進の神経機構を解析した3).
2. 性フェロモンPGF2αを特異的に認識する嗅覚受容体の同定
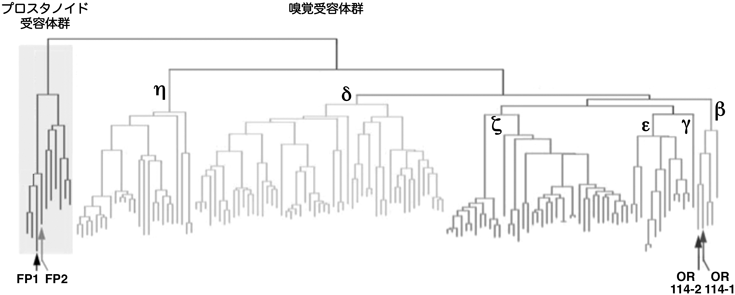
ゼブラフィッシュの嗅上皮には,その形態,分子発現および局在が異なる4種類の嗅細胞(繊毛嗅細胞,微絨毛嗅細胞,Crypt嗅細胞,Kappe嗅細胞)が存在する.まず,PGF2α刺激したゼブラフィッシュの嗅上皮を,神経興奮マーカーである抗リン酸化ERK抗体を用いて免疫組織染色をしたところ,繊毛嗅細胞が特異的に活性化されていることがわかった.各々の繊毛嗅細胞は,約150種類のOR(odorant receptor)ファミリー受容体と約100種類のTAAR(trace amine-associated receptor)ファミリー受容体のうち,どれか1種類を選択的に発現する1).PGF2αはアミノ基を持たない化合物であるため,その受容体が微量アミン関連受容体であるTAARファミリーに属することは考えにくく,ORファミリー受容体に的を絞った.
そこで,ゼブラフィッシュのすべてのOR遺伝子をPCRでクローニングしてcRNAプローブを作製し,PGF2α刺激した嗅上皮における蛍光二重in situハイブリダイゼーション法によって,神経活動マーカーであるc-Fos mRNAの発現と重なりが観察されるOR mRNAを探索した.その結果,PGF2α刺激によってc-Fos陽性となる嗅細胞において嗅覚受容体OR114-1が共発現することがわかった.さらにデータベース検索により,OR114-1と57%のアミノ酸同一性を示す新規のOR遺伝子を発見し,OR114-2と名づけた.これら二つの遺伝子は嗅覚受容体βグループに属し,一般的なプロスタノイド受容体とはきわめて相同性の低いものであった(図1).
次にOR114-1およびOR114-2をHEK293細胞に発現させて,リガンド特異性,濃度依存性を調べた.OR114-1, OR114-2ともに,PGF2αとその代謝産物である15-keto-PGF2αのみによって活性化され,その高いリガンド特異性が明らかとなった.また,OR114-1は高親和性(最小有効濃度:10 nM)),OR114-2は低親和性(最小有効濃度:1 μM)のPGF2α嗅覚受容体であることがわかった.
3. 性フェロモンPGF2αによって活性化される嗅覚神経回路の解析
嗅細胞で受容された匂い・フェロモン情報は,次に嗅覚一次中枢である嗅球の特定の糸球体へと伝えられて匂い地図として表現される.さらにその後,終脳や間脳の高次嗅覚中枢へと情報が伝えられ,匂い地図の解読とともに多様な嗅覚行動の発現へと至る4).そこでまず嗅球において,PGF2αによって活性化される糸球体の同定を試みた.抗リン酸化ERK抗体を用いた嗅球のホールマウント免疫染色およびGCaMP7発現トランスジェニックゼブラフィッシュにおける嗅球のカルシウムイメージングにおいて,PGF2αは腹内側領域の隣接する二つのvmG糸球体を活性化することがわかった.この結果はFriedrichらによって発表されているこれまでの報告5)と合致するものである.
次に,PGF2α刺激によって活性化される高次嗅覚中枢をERKのリン酸化を指標にして解析した.その結果,終脳腹側部腹側核(Vv),視索前核(PPa),外側視床下部(LH)などの領域において,PGF2α刺激に特異的なERKのリン酸化が観察された.
4. PGF2α嗅覚受容体遺伝子欠損ゼブラフィッシュにおける求愛行動の異常
高親和性PGF2α嗅覚受容体OR114-1の生理機能を明らかにするために,TALENゲノム編集技術を用いてOR114-1遺伝子欠損ゼブラフィッシュを作製した.野生型ゼブラフィッシュのオスはPGF2αに対して顕著な誘引行動を示したが,OR114-1欠損体ではそのような行動が観察されなかった.
最後に,野生型メスに対する求愛行動を,野生型オスとOR114-1欠損オスで比較した.野生型オスはメスに対して,追尾(Chase),接触(Touch),回り込み(Encircle)などの求愛行動を頻繁に行ったが,OR114-1欠損オスではこれらの回数・時間が大きく減少していた.以上の結果から,高親和性PGF2α嗅覚受容体OR114-1が,ゼブラフィッシュの求愛行動において重要な役割を果たすことが明らかとなった.
PGF2α, PGD2, PGE2などの各種プロスタグランジンは,それぞれが特異的なプロスタノイド受容体を介して,体内において多様な機能を発現するホルモンあるいはオータコイドとして知られてきた6).本研究では,排卵期のメスの魚から放出されるPGF2αが性フェロモンとして,オスの魚の嗅覚受容体に結合し,求愛行動を引き起こすという個体間コミュニケーションの現象を分子レベル,細胞レベルさらには神経回路レベルで証明し,プロスタグランジン研究の新たな地平を開拓した(図2).
性フェロモンPGF2αによって活性化される高次脳領域として,終脳腹側部腹側核(Vv),視索前核(PPa)が見いだされた.魚のVvは哺乳類の中核野に対応すると考えられており,マウスにおいて中核野が快感や報酬のモチベーションの調節をすることと関連が示唆される7).また,PPaは哺乳類において交尾行動をつかさどると報告されている8).金魚ではVv, PPaを破壊すると産卵行動が著しく阻害され9),サンフィッシュではPPaの電気刺激により,オスの求愛行動や放精が起こる10).以上のように,ゼブラフィッシュで得られた本研究で得られた知見は,これまでに発表されている高次脳領域の生殖行動における機能と合致し,種の壁を越えた性行動の中枢機構の存在が示唆される.
謝辞Acknowledgments
本研究は理化学研究所脳科学総合研究センターシナプス分子機構研究チームのメンバーを中心として,熊本大学杉本幸彦教授のグループ,埼玉大学中井淳一教授のグループと共同で行われたものです.共同研究者の皆様に深く感謝申し上げます.
引用文献References
1) Yoshihara, Y. (2014) in The Olfactory System: from Odor Molecules to Motivational Behaviors (Mori, K. ed), pp. 71–96, Springer, Tokyo.
2) Sorensen, P.W., Hara, T.J., Stacey, N.E., & Goetz, F.W. (1988) Biol. Reprod., 39, 1039–1050.
3) Yabuki, Y., Koide, T., Miyasaka, N., Wakisaka, N., Masuda, M., Ohkura, M., Nakai, J., Tsughe, K., Tsuchiya, S., Sugimoto, Y., & Yoshihara, Y. (2016) Nat. Neurosci., 19, 897–904.
4) Miyasaka, N., Arganda-Carreras, I., Wakisaka, N., Masuda, M., Sumbul, U., Seung, H.S., & Yoshihara, Y. (2014) Nat. Commun., 5, 3639.
5) Friedrich, R.W. & Korsching, S.I. (1988) J. Neurosci., 18, 9977–9988.
6) Narumiya, S., Sugimoto, Y., & Ushikubi, F. (1999) Physiol. Rev., 79, 1193–1226.
7) Olds, J. & Milner, P. (1954) J. Comp. Physiol. Psychol., 47, 419–427.
8) Sakuma, Y. (2008) Ann. N. Y. Acad. Sci., 1129, 55–60.
9) Koyama, Y., Satou, M., Oka, Y., & Ueda, K. (1984) Behav. Neural Biol., 40, 70–86.
10) Demski, L.S., Bauer, D.H., & Gerald, J.W. (1975) J. Exp. Zool., 191, 215–232.
著者紹介Author Profile
 吉原 良浩(よしはら よしひろ)
吉原 良浩(よしはら よしひろ)理化学研究所脳科学総合研究センターチームリーダー.薬学博士.
略歴1961年大阪府に生る.84年京都大学薬学部卒業.89年同大学院薬学研究科博士課程修了.大阪バイオサイエンス研究所,大阪医科大学を経て,98年より現職.
研究テーマと抱負嗅覚入力から行動出力へと至る神経回路の全貌解明.意識を司る脳領域の同定.
ウェブサイトhttp://www.brain.riken.jp/jp/faculty/details/53
趣味読書,スポーツ,将棋.