植物は,光合成によって物質生産を達成している.光合成は,太陽の光エネルギーを利用して大気中のCO2を固定し,炭水化物を生産する一連の反応過程の総称であり,地球上のすべての生命のエネルギー基盤といえる壮大な反応である.ところが,植物が受けている光強度は一定ではなく,時には数秒のオーダーで劇的に変化する.移動能力を持たない植物が,刻一刻と変動する光環境の中で効率よく光合成を行うためには,光合成の場である葉緑体の生理機能を巧みに調節することが必要であろう.実際,葉緑体は,進化の過程でさまざまな光応答戦略を獲得してきた.
その中で我々が注目しているのが,還元力を利用したタンパク質機能の制御系である“レドックス制御”である.この研究分野は,50年以上の長い歴史を持っているが,近年のオミクス解析の進展によるシステム構成因子群の発見を契機として,新たな局面を迎えつつある.本稿では,最近明らかになってきた分子基盤や生理機能に焦点をあてながら,葉緑体レドックス制御システムに関する新たな知見を概説する.
タンパク質のレドックス制御は,酸化還元(reduction-oxidation,レドックス)状態に応じたチオール基の制御(ジスルフィド結合の形成解離など)によって,酵素活性などを調節する分子機構である.レドックス制御の鍵となる役割を担っているのが,還元力伝達因子として働くチオレドキシン(thioredoxin:Trx)である.Trxは分子量約12,000の小さなタンパク質であり,1964年に大腸菌で初めて発見された1).Trxは,WCGPCのアミノ酸配列で構成される活性部位に保存されている一対のシステインペアを用いて,標的タンパク質との間でジチオール/ジスルフィド交換反応を行うことで還元力を伝達する.Trxを介したレドックス制御機構は,原核生物から動植物に至るまで生物界に普遍的に保存されている.
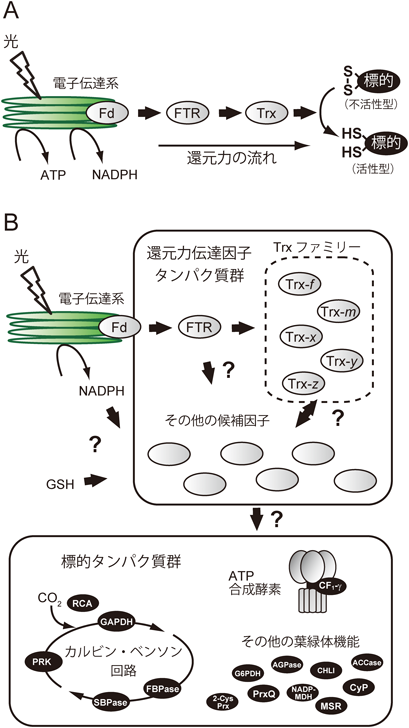
葉緑体のレドックス制御は,光化学反応によって得られる還元力の一部を直接的に用いるために,光に応答した葉緑体の機能調節を可能にするシステムとして理解されている.これまで知られていた還元力経路を図1Aに示す.電子伝達系のフェレドキシンからフェレドキシン-チオレドキシン還元酵素(ferredoxin-thioredoxin reductase:FTR)を介してTrxが還元力を受け取り,次いで還元されたTrxが標的タンパク質を還元する.代表的な標的タンパク質として,カルビン・ベンソン回路中の四つの酵素(GAPDH, FBPase, SBPase, PRK)が知られており,それらは還元によって活性化される.すなわち,この一連の還元力伝達により,炭素固定系を電子伝達系の駆動と連動させることができ,ひいてはより効率的な光合成反応を支えることができる.これらの発見は1970年代にBuchananらによってなしとげられ,それ以来,葉緑体レドックス制御の礎として認識されてきた2).
上述のとおり,(古典的な)システム基盤であるFTR/Trx経路は,単純な一方向の還元力カスケードとして記述されている.ところが,今世紀に入ってゲノミクス・プロテオミクス研究が革新的に進歩したことにより,システム構成因子(還元力伝達因子タンパク質群,および標的タンパク質群)に関する分子カタログ情報が飛躍的に集積された(図1B).それに伴って,“このシステムは,還元力経路を複雑に張り巡らすことで,多様な葉緑体機能の柔軟かつ精緻な調節を達成しているのではないか”というレドックス制御“ネットワーク”の概念が浮上してきた.そして,我々がごく最近得た研究結果は,この仮説を強く裏づけるものであり,従来のシステム像を一変させるものだったのである3).
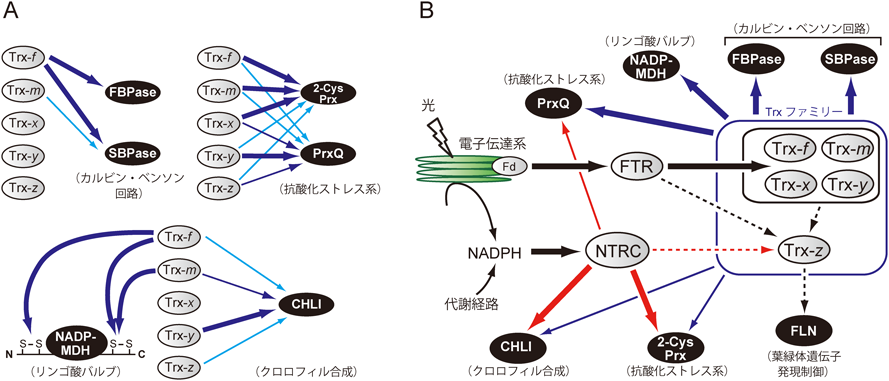
植物ゲノム情報の蓄積によって見つかった還元力伝達因子タンパク質は,実に多岐にわたる.たとえば,葉緑体移行シグナルを持つTrxは,(活性部位以外の)アミノ酸配列の違いを基準として五つのサブタイプに分類される(図1B).我々を含む国内外のグループによって,その機能の多様性を解明しようとする研究が精力的に行われており,たとえば標的タンパク質の選択性の違いなどが部分的に明らかにされた(図2A)4, 5).その他にも,Trxの活性部位と類似した配列を持つ“Trx様タンパク質”など,還元力伝達因子の候補タンパク質をコードする遺伝子も数多く同定されている.しかし,それらのタンパク質の多くは依然として機能未知であり,今後の研究が待たれている.
このようなタンパク質の中で,近年注目を集めているのが,NADPH-チオレドキシン還元酵素C(NADPH-thioredoxin reductase C:NTRC)である6).このタンパク質は,分子構造が独特で,細胞質やミトコンドリアに局在する(いわばオーソドックスな)NADPH-チオレドキシン還元酵素(NADPH-thioredoxin reductase:NTR)のC末端側に,Trxが連結したハイブリッド構造になっている.NTRCは,葉緑体(およびその祖先であるシアノバクテリア)以外には,これまで生物界に見つかっていない.興味深いことに,シロイヌナズナのNTRC欠損変異株は,生育阻害の表現型を示す6, 7).すなわち,NTRCを介した葉緑体のレドックス制御には,植物の生育に何か特別かつ重要な意味があることが想起される.NTRCは,一体どのような機能を持っているのだろうか.2004年の最初の報告以降6),還元力伝達の触媒分子メカニズム解析8)や欠損変異株を用いた生理学解析7, 9, 10)が複数のグループで行われてきたが,その機能に関する統一的な見解は得られていなかった.
我々は,NTRC依存的にレドックス制御を受ける葉緑体機能の全体像を把握するため,まずNTRCの標的タンパク質の探索を行った.この実験では,我々のグループが独自に開発したアフィニティークロマトグラフィー法11)をベースとしたスクリーニングを行い,その後,質量分析を用いて捕捉されたタンパク質を同定した.その結果,複数の葉緑体タンパク質をNTRC標的の候補としてリストアップすることができた.ところが,それらはすべて既知のTrxの標的タンパク質候補と重複しており,当初我々が目論んでいたNTRC特異的かつ新規の標的タンパク質を発見することはできなかった.すなわち,この時点では,NTRCとTrxは共通のタンパク質を標的としているのではないかと考えざるをえなかった.
しかし,ふと思い立って,NTRCとTrx(ここではf型を使用)の標的タンパク質群プロファイルの詳細な量的比較解析を行ったところ,興味深い結果が得られた.標的タンパク質のあるものはNTRCとより多く結合しており,またあるものはTrxとより多く結合していたのである.すなわち,レドックス制御下にある一群の標的タンパク質は,NTRCとTrxに対してそれぞれ異なる親和性を持っていることが強く示唆された.
次に重要なのは,NTRCとTrxに対して異なる親和性を示した標的タンパク質は,実際にNTRCとTrxそれぞれからの還元されやすさも異なるのかどうかを調べることであろう.そこで,該当する標的の組換え体タンパク質を作製し,NTRCと五つのTrxサブタイプからの還元力伝達効率を個別かつ詳細に解析した.その結果,NTRCに対してより多く結合していた標的タンパク質については,NTRCがより高効率な還元力伝達能力を発揮することがわかった.この段階で,次第にNTRCが持つレドックス制御の特性,およびTrxとの違いがみえてきた.その後,タンパク質の組合わせを変えながら解析を続け,NTRCを起点とする還元力ベクトルの向き(標的)と太さ(効率)を順次記述していった(図2B).そして,得られた結果を統合することによって,従来のモデルよりもはるかに高度に組織化された葉緑体レドックス制御ネットワークの実態が浮かび上がってきたのである(図2).
5. FTR/Trx経路とNTRC経路による協調的レドックス制御の意義
ここまでの話を整理しよう.古くから知られていたFTR/Trx経路の他に,NTRC経路という新規の還元力経路が見つかった.しかも,生化学をベースとした実験によって,NTRC経路はFTR/Trx経路とは異なるレドックス制御能を持っていることが明らかになった.もう一つの重要なポイントは,FTR/Trx経路は光による電子伝達系の駆動に絶対的に依存するのに対し,NTRC経路はNADPHを還元力の供給源として用いるため,両者はそれぞれ独立して働くことができることである.では,二つの還元力経路が葉緑体に共存していることは,植物が生きる上でどのような意味を持っているのだろうか.
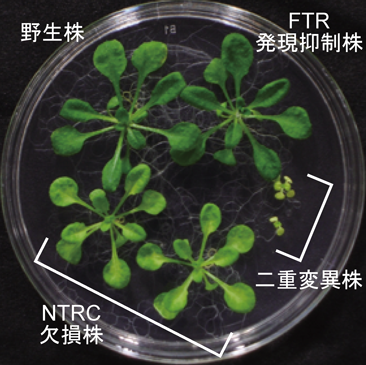
我々は,変異株植物を用いた生理学実験によって,この疑問への回答を得ることを試みた.具体的には,シロイヌナズナを植物材料として,FTR発現抑制株,NTRC欠損株,およびそれらの二重変異株を作出し,その表現型を解析した(図3).それぞれ単独の変異株は,それぞれ異なる生育阻害の表現型を示したものの,両者とも野生株との違いはさほど顕著なものではなかった.しかしながら,二重変異株はきわめて強烈な表現型を示し,独立栄養条件下では発芽後すぐに枯死してしまった.従属栄養条件下では辛うじて生存することができたが(図3),光合成色素量の減少,電子伝達系タンパク質の蓄積量の減少,光合成活性の著しい低下等に示されるように,光合成システム全体が機能不全に陥っていることがわかった.すなわち,FTR/Trx経路とNTRC経路による異なるレドックス制御の協働が,光合成をはじめとする葉緑体の機能調節に重要であり,そして植物の生存戦略に決定的な役割を果たしていたのである3).
本研究の特筆すべき結果として,NTRCがクロロフィル合成や葉緑体遺伝子発現制御に関与するタンパク質に高い親和性を持つことが挙げられる(図2B).さらに,上述したように,NTRCはNADPHを還元力の供給源として用いるため,電子伝達系の光駆動に依存せずに働くことが可能である.これらのNTRCの特性は,葉緑体分化の初期過程における光合成システム構築の制御にとりわけ重要なのかもしれない.実際に,NTRC欠損株の表現型もこの可能性をよく支持している(図3).葉緑体の祖先であるシアノバクテリアでは,NTRCと光合成システム構築に関わる因子とのクロストークは見いだされていない12).以上の結果から,葉緑体が進化の過程で獲得した巧みなレドックス制御の一面が垣間見える.
10年以上前に発見されていたにも関わらず,機能解明が停滞していたNTRCであるが,今回の研究によって,その決定的な役割の一端を解明することができた.この成果は,1970年代にBuchananらが築き上げ,現在の植物生理・生化学の教科書に記述されている葉緑体レドックス制御のシステム基盤を大きく書き換えるものといえる.変異株植物の表現型から推察すれば,FTR/Trx経路とNTRC経路の二つの還元力経路が制御システムの中核であることはほぼ間違いないように思えるが,3節で述べたように,他にも数多くの還元力伝達因子タンパク質がいまだに機能未知のものとして残されている.葉緑体レドックス制御ネットワークの全体像を理解するためには,それらの機能を生化学的に明確にすることが必要であろう.また,多様なシステム構成因子群が,過酷な変動環境を生きる植物体内でそれぞれどのような挙動を示すのか,その機能ダイナミクスを追跡することは,生理学的に非常に意義深い.今後の研究によって,葉緑体レドックス制御ネットワークの構成や振る舞いが,より詳細に明らかになってくるものと期待している.
引用文献References
1) Laurent, T.C., Moore, E.C., & Reichard, P. (1964) J. Biol. Chem., 239, 3436–3444.
2) Buchanan, B.B. (1980) Annu. Rev. Plant Physiol., 31, 341–374.
3) Yoshida, K. & Hisabori, T. (2016) Proc. Natl. Acad. Sci. USA, 113, E3967–E3976.
4) Collin, V., Issakidis-Bourguet, E., Marchand, C., Hirasawa, M., Lancelin, J.M., Knaff, D.B., & Miginiac-Maslow, M. (2003) J. Biol. Chem., 278, 23747–23752.
5) Yoshida, K., Hara, S., & Hisabori, T. (2015) J. Biol. Chem., 290, 14278–14288.
6) Serrato, A.J., Pérez-Ruiz, J.M., Spínola, M.C., & Cejudo, F.J. (2004) J. Biol. Chem., 279, 43821–43827.
7) Pérez-Ruiz, J.M., Spínola, M.C., Kirchsteiger, K., Moreno, J., Sahrawy, M., & Cejudo, F.J. (2006) Plant Cell, 18, 2356–2368.
8) Bernal-Bayard, P., Hervás, M., Cejudo, F.J., & Navarro, J.A. (2012) J. Biol. Chem., 287, 33865–33872.
9) Michalska, J., Zauber, H., Buchanan, B.B., Cejudo, F.J., & Geigenberger, P. (2009) Proc. Natl. Acad. Sci. USA, 106, 9908–9913.
10) Toivola, J., Nikkanen, L., Dahlström, K.M., Salminen, T.A., Lepistö, A., Vignols, H.F., & Rintamäki, E. (2013) Front. Plant Sci., 4, 389.
11) Motohashi, K., Kondoh, A., Stumpp, M.T., & Hisabori, T. (2001) Proc. Natl. Acad. Sci. USA, 98, 11224–11229.
12) Mihara, S., Yoshida, K., Higo, A., & Hisabori, T. (2017) Plant Cell Physiol., 58, 86–94.
著者紹介Author Profile
 吉田 啓亮(よしだ けいすけ)
吉田 啓亮(よしだ けいすけ)東京工業大学科学技術創成研究院化学生命科学研究所助教.博士(理学).
略歴1980年茨城県に生れる.2003年茨城大学理学部卒業.08年大阪大学大学院理学研究科博士課程修了.日本学術振興会特別研究員(PD),東京工業大学資源化学研究所助教を経て,16年より現職.
研究テーマと抱負葉緑体やミトコンドリアなどの植物細胞オルガネラの環境応答,特に酸化還元調節やオルガネラ間クロストークに焦点をあてて研究を行っている.植物の生存戦略の未知なる側面を,分子~個体レベルで縦断的に明らかにしたい.
ウェブサイトhttp://www.res.titech.ac.jp/~junkan/Hisabori_HomePage/index.html
趣味旅行.飲み会.ラーメン.
 久堀 徹(ひさぼり とおる)
久堀 徹(ひさぼり とおる)東京工業大学科学技術創成研究院化学生命科学研究所教授.理学博士.
略歴1957年兵庫県に生れる.80年早稲田大学教育学部卒業.85年同大学院理工学研究科博士後期課程修了.早稲田大学教育学部助手,横浜市立大学文理学部助手,東京工業大学資源化学研究所助教授・教授を経て,16年より現職.
研究テーマと抱負光合成生物のエネルギー変換機構,および,その制御機構に興味をもち,研究手法の開発から生化学的な解析まで,守備範囲を広げて研究を行っている.特に,光合成生物の酸化還元調節機構の多様性と多機能性は,まだ数多くの疑問が残された興味深い研究分野である.
ウェブサイトhttp://www.res.titech.ac.jp/~junkan/Hisabori_HomePage/index.html
趣味ジョギングして脳をリフレッシュすること.