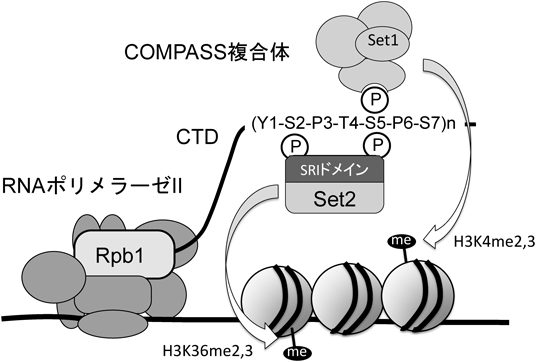
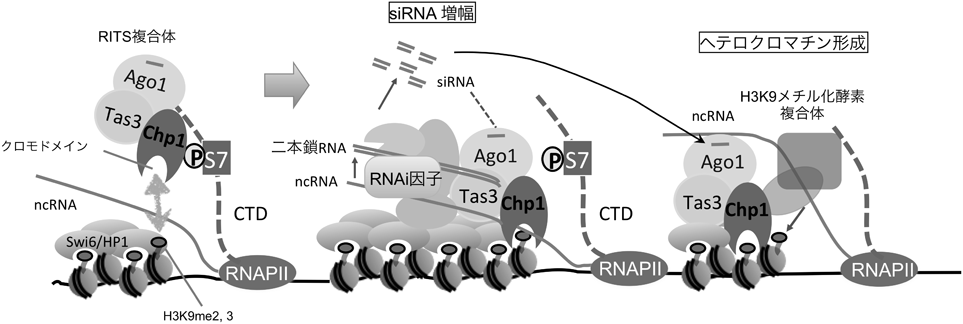
転写装置によるクロマチン構造制御Regulation of Chromatin structure by transcription machinery
北海道大学理学研究院化学部門Department of Chemistry, Faculty of Science, Hokkaido University ◇ 〒060–0810 札幌市北区北10条西8丁目 ◇ N10 W8, Kitaku, Sapporo, Hokkaido 060–0810, Japan
発行日:2017年6月25日Published: June 25, 2017