動物の「細胞外マトリックス」(細胞と細胞の間のスペース)は主に構造タンパク質としてはたらくコラーゲン,コンドロイチン硫酸などの一群の「プロテオグリカン(proteoglycan)」,細胞接着分子などの各種成分で構成されている.一方,植物は細胞の周りが「細胞壁」で囲まれているのが動物細胞との大きな違いの一つである.その主成分は細胞壁多糖であり,その他にタンパク質,糖タンパク質,ならびに本稿で取り上げるプロテオグリカンなどで構成されている.細胞壁多糖は大きくはセルロース,ペクチン,ヘミセルロースの3グループに分けられる.ペクチンやヘミセルロースは細胞壁の分画法(細胞壁からの熱水,キレート試薬,アルカリ試薬による抽出など)に基づく名称で,近年はそれらの局在場所に基づきマトリックス多糖と呼ばれことも多い.このように構成成分は大きく異なるが動物の場合と同じく植物でも「細胞外マトリックス」の用語が一般化している.
植物の細胞外マトリックスにはアラビノガラクタン-プロテイン(arabinogalactan-protein:AGP)と呼ばれる複合糖質が含まれていることが以前から知られており,一般的にはプロテオグリカンに分類されている.動物のプロテオグリカンと似て,AGPの主成分は糖鎖で,少量のコアタンパク質(糖鎖が結合する“芯”となるタンパク質)に結合している.AGPの分子種によってはさらに脂質成分(後述するGPIアンカーのセラミド,図2参照)も含まれている.動物のプロテオグリカンがその主要糖鎖成分としてウロン酸,アミノ糖を含むのとは異なり,AGPは中性糖であるガラクトース(Gal)と植物特有の単糖であるL-アラビノース(L-Ara)が糖鎖の主成分である(本稿では以下断りがない限り糖の構造(光学異性体)を区別するD-は省略してL-は付すことにする).「アラビノガラクタン」の名称もこの2種類の糖に由来している.さらに,動物のプロテオグリカンと同様にAGPもコアタンパク質の分子種は多く,植物組織の分化・成長の制御に関わる多彩な生理機能を持つ情報因子として注目されている.ただし,カラマツ心材のようにコアタンパク質を含まないアラビノガラクタン(AG)も見いだされており,この場合は“多糖”に分類される.また,AGPでもタンパク質含量が高い場合などでは“糖タンパク質(glycoprotein)”と呼ばれることもある.
AGPはその優れた接着・粘着性が着目されて古くから利用されてきた.古代エジプトではミイラを包むときやヒエログリフ(象形文字)を描く天然絵の具としてAGPの一種であるアラビアガム[アカシア(Acacia senegal)の樹液]を使用していたといわれている1).国内では1960年ごろまでは液状のり(アラビアのり)として使われていたが現在は化学合成品が使われている.アラビアガムは現代でも食品,製薬,化粧品などの分野で乳化剤や安定化剤などとして使われている.また,薬用植物に含まれるAGPが免疫系活性化などの生理活性を持つことが知られている.たとえば,和漢生薬の十全大補湯(じゅうぜんたいほとう)に含まれる蒼朮[そうじゅつ,キク科のホソバオケラ(Atractylodes lancea)の根茎]のAGP糖鎖はT細胞を活性化して骨髄細胞の増殖をもたらす2).本稿ではAGPの特性,構造,合成と分解,生理機能などについて我々の知見も含めて紹介する.
2. アラビノガラクタン-プロテイン(AGP)とは?
1)AGPの分布,コアタンパク質と糖鎖
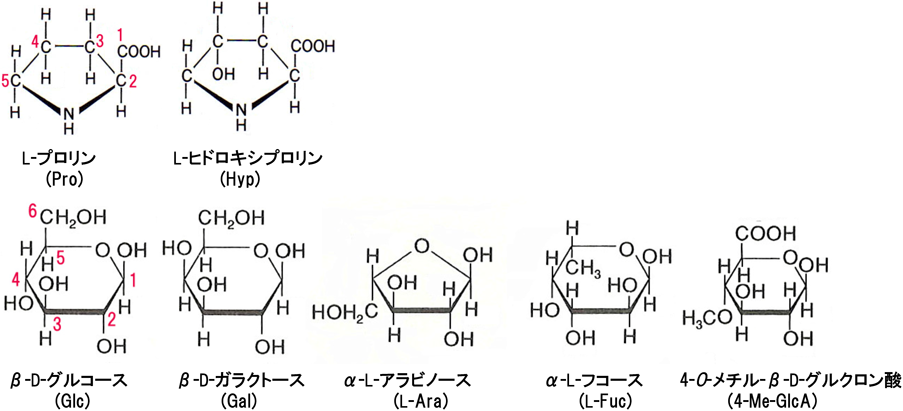
AGPの特徴の一つは,被子植物や裸子植物,コケ植物,さらには藻類[例,単細胞緑藻類のクロレラ(Chlorella)の熱水抽出物]などのあらゆる植物の器官・組織・細胞に普遍的に存在していることである.植物細胞の懸濁培養によっても培地中に分泌される.しかしながらその含量はペクチンやヘミセルロースなどの細胞壁多糖と比べてはるかに少なく,一般的には植物組織乾燥重量の1%以下である.我々がダイコン根からAGPを抽出精製したところでは生重量1 kg(ダイコン成根1本相当)から得られる精製AGPは100 mg程度であった.一般に,AGPは糖含量が高く(≧90%),コアタンパク質は全体の約10%以下である.分子種によって異なるが,コアタンパク質にはヒドロキシプロリン(Hyp,図1)が多く含まれる.植物細胞壁にはAGPの他にもHypに富む成分が含まれており,Pro-rich protein(PRP),エクステンシン(extensin)と呼ばれる.AGPに多く含まれるその他のアミノ酸残基としてはプロリン(Pro),アラニン(Ala),セリン(Ser),トレオニン(Thr)などである.AGPの糖鎖は上述したようにGalとL-Araに富む.AGPの分子種によってはさらに,グルクロン酸(GlcA)または4-O-メチル-グルクロン酸(4-Me-GlcA),L-フコース(L-Fuc),キシロース(Xyl)などの糖残基が含まれる(図1)3).
2)AGPの生合成過程
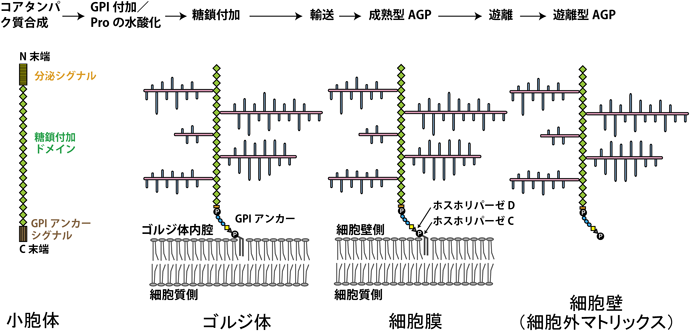
AGPの合成過程は基本的には動物のプロテオグリカンの合成過程とほぼ同様であり,コアタンパク質は植物でも小胞体で合成され,次にゴルジ体へ運ばれる.この過程で,合成されたコアタンパク質は翻訳後修飾(post-translational modification)を受け,N末端の分泌シグナルが除かれ,Proが酵素によって水酸化されてHypが生じる(図2).次に,主にゴルジ体内でさまざまな糖転移酵素(糖鎖合成酵素)の働きでAG糖鎖が付加される.糖鎖が付加されるアミノ酸残基は動物のプロテオグリカンとは異なり,AGPの場合は一部のHyp残基のO-4位水酸基に糖が逐次転移される.
一方,コアタンパク質のC末端にはグリコシルホスファチジルイノシトール(glycosylphosphatidylinositol:GPI)アンカーシグナルが付加しており,GPIアンカーがコアタンパク質に結合することでAGP分子が小胞体とゴルジ体の膜につなぎ止められる4).その後,小胞輸送によってAGPは細胞膜まで運ばれる.ホスホリパーゼCまたはDの働きでGPIアンカーが切り離されて,AGPは水可溶性成分として細胞壁(細部外マトリックス)に遊離されると考えられている1, 4–8).模式図(図2)はAGP分子種の「クラシカル(classical)AGP」についての合成過程である.その他に,リシン(Lys)に富むドメインを持つコアタンパク質,Hypに富む糖鎖付加シグナル以外にファシクリン様(fasciclin-like)ドメインを持つコアタンパク質,ペプチド鎖が短いAGP(AG peptide),GPIドメインを持たないAGP(non-classical AGP Lys)などが知られており,模式図とは異なる遊離型AGPが生じる.
AGPの代謝回転(turnover)は他の細胞壁多糖と比べて大変速く,合成されたAGPの33%が1時間で分解される9).分解で生じたアミノ酸,単糖の一部は細胞内に取り込まれて再利用されると考えられる.単糖は動物の場合と基本的には同じ再利用経路(後述)と呼ばれる代謝経路で「糖ヌクレオチド」に変換される.各種糖ヌクレオチドが出発材料となってAGP糖鎖がリサイクル合成される.
3)AGP分子の多様性
AGPの特徴としてさらに器官・組織特異的発現があげられる.AGPのアミノ酸組成,糖成分と糖鎖構造は植物の器官(花,葉,根など)の違いにより異なり,また,植物の成長に伴い変化することが知られている.たとえば,ダイコンの場合,若い根(スーパーマーケットで売られているカイワレダイコン程度)のAGPは糖鎖にL-Fuc残基を含む.L-Fucは動物の血液型O(H)型の抗原決定基であり,ダイコンでも同様な血液型類似活性を示す.しかし,植物が成長して根が肥大するとL-Fucは消失し,代わりに成葉AGPにL-Fucが見いだされるようになる10).AGPのコアタンパク質は多種類存在し,シロイヌナズナでは,AGPのコアタンパク質をコードする遺伝子は80以上も存在する.これらの遺伝子が成長段階と各種器官で特異的に発現されてさまざまな糖鎖構造を持つAGP分子種が生じると考えられている8).AGPは分化・成長の制御に関わる多彩な生理機能を持つ情報因子として注目されている1, 5–8).しかしながら,生理機能にはコアタンパク質と糖鎖のどちらが重要なのか,糖鎖が重要であるとしたらその糖鎖構造は何か,などについては不明な点が多い.
4)AGP特異的な結合試薬
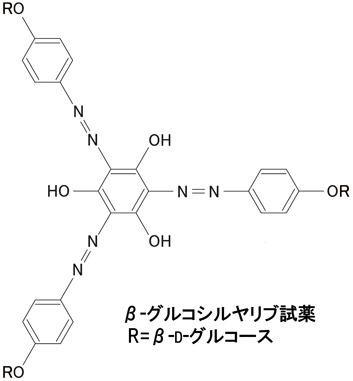
AGPに特異的に結合するヤリブ試薬[Yariv reagent,化学名1,3,5-tri(p-glycosyloxyphenylazo)-2,4,6-trihydroxybenzene]が1962年にYariv博士らによって化学合成された(図3)11).ヤリブ試薬はAGPと特異的に反応して複合体(赤色の沈殿)を生じるので,組織からのAGPの精製,定量,AGPの生理機能解析(後述),等に用いられてきた.ヤリブ試薬のエピトープ(ヤリブ試薬が結合するAGPの部位・構造)はヤリブ試薬の開発以来,約50年間不明だったが,我々はヤリブ試薬が後述するAGP糖鎖の主鎖であるβ-(1→3)-ガラクタンを認識して結合することを明らかにした12).今までにAGP糖鎖構造を持っていてもヤリブ試薬と反応しないAGPやAGが知られているが,β-(1→3)-ガラクタンに他の糖が多数結合してヤリブ試薬結合部位が“マスク”されているためと思われる.また,他の植物多糖の研究と同様に,AGPの局在性や生理機能を調べるツールとしてさまざまなAGP糖鎖認識抗体が作成されている6).
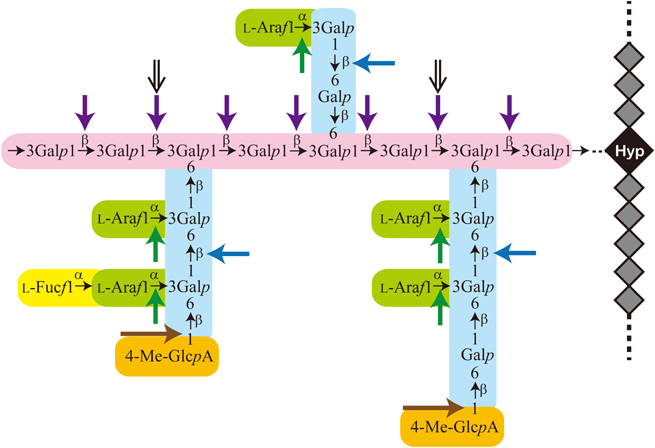
上述のようにAGPの糖鎖は主にGalとL-Araで構成され,さらにその他の糖が結合している.AGP糖鎖の基本構造は連続したβ-(1→3)-ガラクトシル残基からなる主鎖に,β-(1→6)-ガラクトシル残基からなる側鎖が分岐結合したβ-(1→3)/(1→6)-ガラクタンである.このガラクタン骨格の側鎖にはL-Ara残基が分岐結合している(図4).ダイコン成根AGPの場合,側鎖の非還元末端にGlcAまたは4-Me-GlcAが結合している.また,ダイコン成葉ではL-Ara残基の先にL-Fucが結合する3, 13).植物細胞壁多糖は一般的に複雑な構造であるが,その中でもAGPの糖鎖はかなり複雑である.そのため,糖鎖–生理機能の相関性の解析はなかなか進展しなかった.
糖鎖の構造解析にはさまざまな分析手法が用いられるが,酵素による特異的断片化も糖鎖の構造解析,糖鎖の生理機能解析に有効である.我々がAGPの研究を始めたころは糖鎖の分解酵素としてはL-Ara残基に作用するα-L-アラビノフラノシダーゼ以外は明確には知られていなかった.AGPの特性である組織中の含量の少なさ,糖鎖構造の複雑性がAGP糖鎖の分解酵素活性探索・酵素精製の足かせになっていたためと思われる.我々は,酵素活性測定に適した基質調製から始めて,酵素の探索,精製,遺伝子クローニングを行ってきた.今までにエキソ-β-(1→3)-ガラクタナーゼ,エンド-β-(1→6)-ガラクタナーゼ,エンド-β-(1→3)-ガラクタナーゼ,β-グルクロニダーゼ,等を見いだしてその性質とAGP糖鎖への作用様式を明らかにしてきた.表1に微生物,植物から精製・遺伝子クローニングされているAGP糖鎖分解酵素一覧を示す.表では起源が異なる酵素1, 2例のみを取り上げた.さらに,各種酵素の作用点のまとめを図4の糖鎖構造模式図に重ねて示す.AGP糖鎖分解酵素については総説・解説14–16)で紹介されている.
表1 AGP糖鎖の分解酵素| 酵素名 | EC番号a) | GHファミリーb) | 起源 | 文献 |
|---|
| エキソ-β-(1→3)-ガラクタナーゼ | 3.2.1.145 | GH43 | ウスバタケ(Irpex lacteus) | 17 |
| エンド-β-(1→3)-ガラクタナーゼ | 3.2.1.181 | GH16 | エノキタケ(Flammulina velutipes) | 18 |
| エンド-β-(1→6)-ガラクタナーゼ | 3.2.1.164 | GH30 | アカパンカビ(Neurospora crassa) | 19 |
| β-ガラクトシダーゼ* | 3.2.1.23 | GH35 | 子嚢菌(Hypocrea jecorina) | 20 |
| ダイコン(Raphanus sativus) | 21 |
| α-L-アラビノフラノシダーゼ* | 3.2.1.55 | GH3, 43, 54 | 赤色酵母(Rhodotorula flava) | 22 |
| ダイコン(Raphanus sativus) | 23 |
| β-L-アラビノピラノシダーゼ | 3.2.1.88 | GH27 | 放線菌(Streptomyces avermitilis) | 24 |
| α-L-ラムノシダーゼ | 3.2.1.40 | GH78 | 放線菌(Streptomyces avermitilis) | 25 |
| α-(1→2)-L-フコシダーゼ | 3.2.1.63 | (GH29, 95)c) | 細菌(Xanthomonas manihotis) | 26 |
| β-グルクロニダーゼ* | 3.2.1.31 | GH79 | クロコウジカビ(Aspergillus niger) | 27 |
| シロイヌナズナ(Arabidopsis thaliana) | 28 |
| a)EC番号(酵素番号,Enzyme Commission numbers)は酵素反応の種類と基質特異性の違いによる酵素の分類法である.EC3.2.1.xは糖質加水分解酵素を,xは酵素の基質特異性を表す.b)GHファミリーはCAZy(carbohydrate-active enzyme, http://www.cazy.org/)による酵素のアミノ酸配列に基づく分類法である.GHは糖質加水分解酵素(glycoside hydrolase)を表し,現在(2017年)は135のファミリーに分類されている.c)Xanthomonasの酵素(市販品)はGHファミリー未分類.カッコ内はα-L-フコシダーゼの既知GHファミリー.*微生物の他に植物でも見いだされている酵素を示す. |
1)微生物起源の酵素
エキソ-β-(1→3)-ガラクタナーゼはキノコの仲間のウスバタケ(Irpex lacteus)由来の酵素製剤「ドリセラーゼ」(協和発酵バイオ(株)製,当時)に活性を認めた.キノコは木材腐朽菌として,植物組織を分解して栄養源としているので,各種の植物細胞壁多糖分解酵素の宝庫といえる.本酵素は図4のようにAGP糖鎖のβ-(1→3)-ガラクタン主鎖の非還元末端から糖鎖を順番に切り出す.β-(1→6)-ガラクタン側鎖が結合している分岐部分はバイパスして分解が進行するので,側鎖が結合していない場合はGalが遊離し,側鎖はオリゴ糖として丸ごと切り出される17, 29).本酵素はAGPの糖鎖構造解析に有用なツールである3, 13).一方,エンド-β-(1→3)-ガラクタナーゼはAGP糖鎖のβ-(1→3)-ガラクタン主鎖の内部に作用してオリゴ糖を生じる酵素である18).エンド-β-(1→6)-ガラクタナーゼはAGP糖鎖のβ-(1→6)-ガラクタン側鎖に作用してGalとβ-(1→6)-ガラクトオリゴ糖を生じる19).β-グルクロニダーゼは以前から知られている酵素であるが,我々が見いだしたのはAGP糖鎖の非還元末端に結合しているGlcAと4-Me-GlcA残基の両方に作用する酵素である27).AGP分子種によってはさらにL-Fuc, Xyl,L-アラビノピラノース残基などが含まれており,それらに作用する酵素が見いだされている(表1).このように,AGP糖鎖のβ-(1→3)/(1→6)-ガラクタン骨格へ作用する酵素を用いた糖鎖の特異的断片化と各種グリコシダーゼによる糖鎖のトリミングが可能となってきた.これらの酵素を適用した糖鎖の構造解析と生理機能解析の進展が期待される.
2)植物体内でのAGP糖鎖の分解
AGPは他の細胞壁多糖と比べて代謝回転が速い.また,糖鎖構造が複雑なのでAGP糖鎖の植物体内での分解には細胞壁で働く複数種の酵素が必要となる.植物にはエキソ-β-(1→3)-ガラクタナーゼのような多糖分解酵素(glycanase)ははっきりとは確認されておらず,低重合度のオリゴ糖に主に作用するβ-ガラクシダーゼやα-L-アラビノフラノシダーゼなどのグリコシダーゼ(glycosidase)の協同作用でAGP糖鎖の分解が進行すると考えられる.実際に,ダイコンのAGP糖鎖にダイコン由来のβ-ガラクトシダーゼを単独で作用させても糖鎖はあまり分解されないが,α-L-アラビノフラノシダーゼとβ-グルクロニダーゼとともに働かせると糖鎖の90%以上が分解される30).酵素作用で生じた単糖の一部は細胞内に取り込まれて再利用経路に入ると考えられる.
一方,AGPコアタンパク質もタンパク質分解酵素の働きですばやく分解され,生じたアミノ酸の一部は糖の場合と同じく再利用されると考えられる.AGPコアタンパク質はHypやProに富んでいるので特異的タンパク質分解酵素の存在が推定される.我々は最近,エノキタケ培養液にHyp残基に特異性が高いプロテアーゼ/ペプチダーゼを見いだし,AGPコアタンパク質を分解することを明らかにした31).しかしながら,植物体内で働くタンパク質分解酵素については不明のままである.
AGPの糖鎖に限らず,植物細胞壁の糖鎖の大半は,各種糖ヌクレオチドから糖転移酵素(糖鎖合成酵素)の働きで合成される.高度に分岐し,多様な末端糖残基を持つAGPの糖鎖の合成には,多数の酵素が関わると考えられる.糖転移酵素の多くはきわめて基質特異性が高い.そのため,同じグリコシルドナー(たとえば,Gal残基)の転移であっても,結合が異なる糖残基[たとえば,β-(1→3)-結合のGal残基とβ-(1→6)-結合のGal残基]の転移には,別の糖転移酵素が働くのが常である.さらに,同じ結合の同じ糖残基であっても,グリコシルアクセプター(糖を転移する相手)が異なれば,違う糖転移酵素が働く場合も多い.したがって,AGP糖鎖の基本骨格であるβ-(1→3)/(1→6)-ガラクタンの合成では,コアタンパク質のHyp残基に最初のGalを転移する酵素,β-(1→3)-ガラクタン主鎖を伸長する酵素,β-(1→3)-ガラクタン主鎖に分岐を作る酵素,側鎖のβ-(1→6)-ガラクタンを伸長する酵素は,それぞれ異なると考えられる.これらに加えて,AGPの糖鎖末端には,α-L-Araやβ-GlcA(または4-Me-β-GlcA),α-L-Fucなどの糖が結合し,それぞれに複数の存在様式(結合様式と結合相手)が存在することを考えると,AGP糖鎖の合成には10種類以上の糖転移酵素が関わっていると予想される.
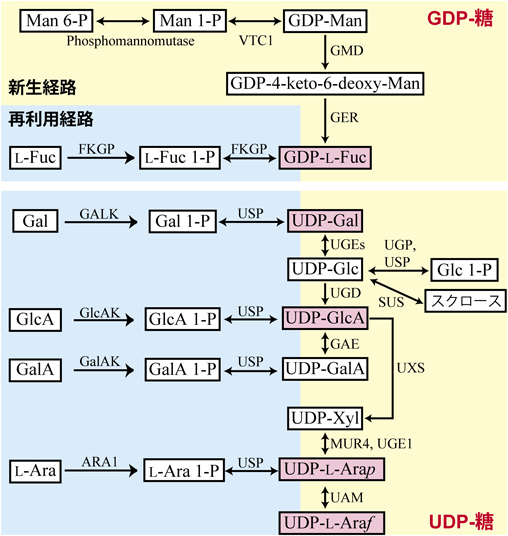
1)糖ヌクレオチドの合成経路
細胞壁の糖鎖の大半は,糖ヌクレオチドであるウリジン5′-二リン酸(UDP)-糖やグアノシン5′-二リン酸(GDP)-糖が出発材料(グリコシルドナー)となって糖転移酵素の働きで合成される.主要な糖ヌクレオチドはUDP-GalとUDP-L-Ara, UDP-GlcA, GDP-L-Fucなどである.植物ではUDP-糖代謝経路の初発物質は,UDP-Glcである.UDP-Glcは,グルコース1-リン酸(Glc 1-P)やスクロースから合成される(図5).UDP-Glcは,GlcのC-6位が酸化されるとUDP-GlcAとなる.UDP-GlcAはさらにC-6位の脱炭酸反応によりUDP-Xylとなる.ここまでのUDP-糖代謝経路は種間で高度に保存されており,我々ヒトを含む哺乳類でも同じ機構が存在する.しかし,植物ではこの先の経路が異なっている.植物では,UDP-XylがC-4エピマー化されUDP-L-Araとなる.AGPの主要構成糖であるL-Araはこの反応により合成される32).L-Araは一部の限られた動物にしかみられず,UDP-L-Ara合成経路は動物ではほとんど存在しない.
植物では,UDP-GlcからUDP-L-Araまでの経路が,ゴルジ体と細胞質基質の両方に存在する.つまり,細胞質基質で合成されたUDP-GlcAの一部(または大半)は,UDP-GlcA輸送体によりゴルジ体内腔に輸送され,細胞質基質とは異なる酵素によりUDP-XylやUDP-L-Araに変換されている.糖転移酵素の多くは,触媒部位はゴルジ体内腔にあるため,ゴルジ体にこのようなUDP-糖合成経路が存在し,UDP-L-Araを糖転移反応の基質として利用しているのは合目的といえる.一方で,細胞質基質で合成されるUDP-L-Araも別の輸送体によりゴルジ体内腔に輸送されると考えられており,AGPの糖鎖も含めてL-Araを含む細胞壁の糖鎖の合成には,ゴルジ体経路と細胞質基質経路の両方が貢献していると考えられる.
植物細胞壁の糖鎖に含まれるL-Ara残基の環状構造に関しては大きな謎があった.UDP-Xylから合成されるUDP-L-AraのL-Ara部分はピラノース型(六員環)であるのに,AGPに限らず,細胞壁の糖鎖にみられるL-Ara残基の大半はフラノース型(五員環,図1参照)なのである.今から10年ほど前,UDP-L-Araのピラノース型(UDP-L-Arap)をフラノース型(UDP-L-Araf)に変換するUDP-L-アラビノピラノースムターゼ(UAM)が発見された33).この酵素は細胞質基質でこの反応を触媒していることがわかった.しかしながら,ゴルジ体で合成されたUDP-L-Araはどのように細胞質基質のUAMの反応を受け,またゴルジ体に戻ってくるのだろうか? この謎はまだ解かれていないが,ゴルジ体から細胞質基質にUDP-L-Arapを輸送する輸送体と,細胞質基質からゴルジ体にUDP-L-Arafを輸送する未知の二つの輸送体が存在する可能性が高い.
2)糖転移酵素
AGPのコアタンパク質は小胞体で合成され,Pro残基はプロリン4-ヒドロキシラーゼの作用を受けて水酸化されHypになる(図1, 2).この過程は動物のコラーゲンに含まれるHypが生じる反応と基本的には同じ反応である.コアタンパク質にはさらにさまざまな糖転移酵素の働きで,Hypが非連続的に並んでいる配列(Ala-HypやSer-Hypの繰り返し配列)にAG糖鎖が付加されると考えられる(Hyp-contiguity hypothesis).一方,連続したHypモチーフはAGP分子種によってはアラビノオリゴ糖で修飾されると予測される34).AGP糖鎖の合成に関わる主要な糖転移酵素を表2にまとめた.糖鎖合成の最初の反応は,Hyp残基へのGalの付加である.この反応を触媒する酵素として2種類の糖転移酵素が同定されている.Golgi-localized hydroxyproline-O-galactosyltransferase(GALT)とhydroxyproline galactosyltransferase(HPGT)はどちらも糖転移酵素ファミリー(GTファミリー,表1のGHファミリー同様に糖転移酵素はおよそ100のファミリーに分類されている)31に属するが,GALTは700アミノ酸残基程度のタンパク質であるのに対して,HPGTは340アミノ酸残基程度と,そのサイズには大きな違いがある.両酵素ともHyp残基にGalを転移する活性が検出されており,生体内で同じ反応を触媒している可能性が高い.現段階では,多種類のコアタンパク質のどれに優先的に働くかなどの両酵素の特異性の違いや生理的役割の違いなどは不明である.
表2 AGP糖鎖の合成に関わる糖転移酵素| 酵素名 | GTファミリー | 基質糖ヌクレオチド | 触媒する反応 | 文献 |
|---|
| Hyp-O-ガラクトシルトランスフェラーゼ | GT31 | UDP-Gal | コアタンパク質への最初のGalの付加 | 35, 36 |
| β-(1→3)-ガラクトシルトランスフェラーゼ | GT31 | UDP-Gal | β-(1→3)-ガラクタンの伸長 | 37 |
| β-(1→6)-ガラクトシルトランスフェラーゼ(分枝形成および側鎖伸長) | GT29 | UDP-Gal | β-(1→6)-ガラトシル側鎖の形成とβ-(1→6)-ガラクタン側鎖の伸長 | 38 |
| β-(1→6)-ガラクトシルトランスフェラーゼ(側鎖伸長) | GT31 | UDP-Gal | β-(1→6)-ガラクタン側鎖の伸長 | 39 |
| β-グルクロノシルトランスフェラーゼ | GT14 | UDP-GlcA | β-(1→6)-ガラクタン側鎖末端へのGlcAの付加 | 40, 41 |
| α-L-アラビノフラノシルトランスフェラーゼ | 未同定 | UDP-L-Araf(予想) | β-(1→6)-ガラクタン側鎖へのL-Arafの付加 | — |
| α-(1→2)-L-フコシルトランスフェラーゼ | GT37 | GDP-L-Fuc | 糖鎖末端L-Ara残基へのL-Fucの付加 | 26, 42, 43 |
β-(1→3)-ガラクタン主鎖の伸長は,β-1,3-ガラクトシルトランスフェラーゼによって触媒される.この酵素もGTファミリー31に属している.β-(1→3)-ガラクタン主鎖にβ-(1→6)-結合でGalの分岐を形成する酵素は,GT29のβ-(1→6)-ガラクトシルトランスフェラーゼである.しかしながら,この分岐からβ-(1→6)-ガラクタン側鎖を伸長する酵素は,同じ酵素が触媒する可能性とGT31ファミリーの別の酵素が触媒する可能性が指摘されている.
β-(1→6)-ガラクタン側鎖にGlcAやL-Fucを付加する糖転移酵素として,それぞれGT14ファミリーのβ-グルクロノシルトランスフェラーゼ(GlcAT14)とGT37ファミリーのα-(1→2)-L-フコシルトランスフェラーゼ4(FUT4)およびFUT6が同定されている.L-Araの付加に関わるα-L-アラビノフラノシルトランスフェラーゼは未同定である(表2).また,表には示さないが,一部のAGP糖鎖にはXyl残基やL-ラムノース(L-Rha)残基も見つかっており,これらはそれぞれUDP-XylとUDP-L-Rhaから合成されると考えられる.
動物のプロテオグリカンを構成するコンドロイチン硫酸やヘパラン硫酸が動物にとって生理的にきわめて重要な分子であるのと同様に,植物のプロテオグリカンであるAGPも植物のさまざまな生命現象に関わっている.AGPは重量の大半を糖鎖が占めており,糖鎖は分子機能にきわめて重要と予想されるが,AGP糖鎖の生理活性を示した研究例は限られている.
ゲノムサイズの小さいシロイヌナズナでも,AGPのコアタンパク質をコードする遺伝子は80以上も存在する.分子種間には機能的な重複があり,単一AGP遺伝子の欠損では表現形質が変化しにくいことが,AGPの機能解析を難しくしている.さらに,AGP糖鎖構造が複雑で不均質(heterogeneity)であることが糖鎖–機能相関性解析を困難にしている.
1)情報分子としての働き
1995年に権威ある雑誌『Cell』に,植物生殖におけるAGPの機能に関する2報の論文が発表された44, 45).米国の研究グループによるこれらの論文では,タバコの花柱のtransmitting tissue-specific protein(TTSタンパク質)と呼ばれるAGPが花粉管を受精へと導くこと(ガイダンス)が示された.TTSタンパク質が花粉管を誘引することや,花粉管によりAG糖鎖の分解を受けることも併せて示された.大変興味深い現象であるが,TTSタンパク質の糖鎖構造や生理活性に関する研究がなく,そのメカニズムは解明されなかった.この研究から20年後,我々はまったく別のアプローチで,AGP糖鎖の側鎖構造が花粉管ガイダンスに関わることを示す研究に参画できた46).名古屋大学のグループは,2009年に,雌性配偶体の助細胞から分泌される花粉管誘引ペプチド,LUREを発見していた47).しかしながら,花粉管がLUREに応答するには,未同定の雌しべ側因子,activation molecule for response capability(AMOR)と先に接触する必要があることがわかっていた.AMORの精製や同定から,AMORの実体はAGP糖鎖の側鎖にみられる,4-Me-β-GlcA-(1→6)-Galという構造(図1参照)であることが判明した46).興味深いことに,メチル基がないβ-GlcA-(1→6)-Galやアノマー構造である4-Me-α-GlcA-(1→6)-GalはほとんどAMOR活性を持たない.このことは,花粉管が4-Me-β-GlcA-(1→6)-Galを厳密に認識していることを示唆している.我々が知る限り,AGPの特定の糖鎖部位・配列が植物の生命現象に関わることが示された唯一の例である.TTSタンパク質とは異なる花粉ガイダンス現象を観察しているため,AMORとTTSが同一分子かはわからないが,4-Me-β-GlcA-(1→6)-GalはAGP糖鎖によくみられる構造である.
ヒャクニチソウ培養細胞から単離されたxylogenと呼ばれるAGPも,AGPが細胞分化に情報分子として関与することを示す好例である48).xylogenは1か所または2か所がAG糖鎖で修飾されている非特異的脂質輸送タンパク質であり,培養細胞の管状要素(導管細胞など)への分化を誘導する活性を持つ.xylogenタンパク質は典型的なAGPではないが,化学処理により糖鎖を除去するとその活性を失うため,AG糖鎖がその機能に重要であると考えられる.ヤリブ試薬(図3)処理でヒャクニチソウの分化率が著しく低下することも,AGPの関与を示している.これらの事実は,AG糖鎖が情報分子の活性に重要であることを示唆している.植物には,特定のAG糖鎖を認識する未知の結合(受容体)タンパク質と,その働きを介した情報伝達の機構が存在する可能性がある.
2)分化や成長における働き
我々は,AGPにはAMORのような情報分子としての働きに加えて,細胞表層プロテオグリカンとして細胞成長や細胞形態を制御する働きがあると考えている.分子機能を解明する上で,シロイヌナズナやイネの突然変異体の解析は,大変有効なアプローチであるが,AGPは分子種が多く分子種間で機能が重複しているため,突然変異体を利用した研究例は限られている.
シロイヌナズナのsalt overly sensitive 5(sos5)は,細胞接着ドメインであるファシクリン様ドメインを持つAGP分子種,fasciclin-like AGP 4(FLA4)を欠損した突然変異体である49).sos5は,その名前のとおり,NaClを含む培地で根の成長遅延を示す突然変異体として単離された.この突然変異体では,根の先端で組織形態が乱れており,このことがNaClへの耐性低下を引き起こしていると考えられている.一方で,この形質は,FLA4が根の先端の細胞接着や細胞形態の制御に関わることを強く示唆している.その他のAGP分子種も植物の分化・成長に共通する生理的な働き,ないしはAGP分子種ごとの固有な働きを担っていると考えられる.
AGPの特異的な阻害剤であるヤリブ試薬(図3)を用いた研究でも,AGPが細胞形態の制御に関わることが示唆されている.タバコの培養細胞をヤリブ試薬で処理すると,水風船のように一部が異常に膨れるなど細胞の形態異常が生じる.これらの表層微小管を観察すると,もともと細胞の長軸に対して水平方向に配向していた表層微小管の配向が乱されていることが報告されている50).ヤリブ試薬による表層微小管の変化は,生きたシロイヌナズナの根でも観察されている51).これらの実験結果は,細胞外に存在するAGPと細胞内の表層微小管との間に何らかの連絡があることを示唆している.
今後のAGP研究では,AG糖鎖の分子機能が間違いなく興味の中心となる.AG糖鎖に特異的に結合するヤリブ試薬が,植物のさまざまな生命現象を阻害・攪乱することは,AG糖鎖が何らかの重要な働きをしていること示唆している.しかしながら,“重要な働き”は複数あると我々は考えている.AMORやxylogenの研究では,AG糖鎖が情報分子として働くことが示された.一方で,前述のヤリブ試薬による表層微小管の配向変化は,情報分子としての働きとは別に,AGPが細胞外の変化を細胞内部に伝達する働きを持つことを示唆している.AGPと表層微小管との間にどのような連絡があるかは,AGP研究者の今後の最大の興味の一つである.
AGP研究の大きな障壁の一つが,分子種の多さと糖鎖構造の複雑さである.一つのAGP分子種の欠損により幸運にも突然変異体で形質変化を観察できた例もあるが,多くのAGPは機能が互いに重複しており,分子種の多さから多重変異体の解析も容易でない.そこで我々は,植物生体内でAG糖鎖の破壊を起こす実験系の構築を進めている.AG糖鎖は不均質で多様な構造をとるが,糖鎖主鎖がβ-(1→3)-ガラクタンである点は共通している.真菌由来のエキソ-β-(1→3)-ガラクタナーゼを植物生体内で発現させることで,分子種を問わずにAGPの機能不全を起こすことができると期待される.
AGPは植物特有な分子であり,かつ陸上植物では高度に保存された分子でもある.今後も,AGPの分子機能解析を通して,植物の生命現象を解明していきたい.
引用文献References
1) Majewska-Sawka, A. & Nothnagel, E.A. (2000) Plant Physiol., 122, 3–9.
2) Yu, K.W., Kiyohara, H., Matsumoto, T., Yang, H.C., & Yamada, H. (1998) Planta Med., 64, 714–719.
3) Inaba, M., Maruyama, T., Yoshimi, Y., Kotake, T., Matsuoka, K., Koyama, T., Tryfona, T., Dupree, P., & Tsumuraya, Y. (2015) Carbohydr. Res., 415, 1–11.
4) Oxley, D. & Bacic, A. (1999) Proc. Natl. Acad. Sci. USA, 96, 14246–14251.
5) Gaspar, Y., Johnson, K.L., McKenna, J.A., Bacic, A., & Schultz, C.J. (2001) Plant Mol. Biol., 47, 161–176.
6) Showalter, A.M. (2001) Cell. Mol. Life Sci., 58, 1399–1417.
7) Seifert, G. & Roberts, K. (2007) Annu. Rev. Plant Biol., 58, 137–161.
8) Ellis, M., Egelund, J., Schultz, C.J., & Bacic, A. (2010) Plant Physiol., 153, 403–419.
9) Gibeaut, D.M. & Carpita, N.C. (1991) Plant Physiol., 97, 551–561.
10) Tsumuraya, Y., Ogura, K., Hashimoto, Y., Mukoyama, H., & Yamamoto, S. (1988) Plant Physiol., 86, 155–160.
11) Yariv, J., Rapport, M.M., & Graf, L. (1962) Biochem. J., 85, 383–388.
12) Kitazawa, K., Tryfona, T., Yoshimi, Y., Hayashi, Y., Kawauchi, S., Antonov, L., Tanaka, H., Takahashi, T., Kaneko, S., Dupree, P., Tsumuraya, Y., & Kotake, T. (2013) Plant Physiol., 161, 1117–1126.
13) Haque, M.A., Kotake, T., & Tsumuraya, Y. (2005) Biosci. Biotechnol. Biochem., 69, 2170–2177.
14) 一ノ瀬仁美,金子 哲(2011)応用糖質科学,1, 232–237.
15) Sakamoto, T. & Ishimaru, M. (2013) Appl. Microbiol. Biotechnol., 97, 5201–5213.
16) Knoch, E., Dilokpimol, A., & Geshi, N. (2014) Front. Plant Sci., 5, 198.
17) Kotake, T., Kitazawa, K., Takata, R., Okabe, K., Ichinose, H., Kaneko, S., & Tsumuraya, Y. (2009) Biosci. Biotechnol. Biochem., 73, 2303–2309.
18) Kotake, T., Hirata, N., Degi, Y., Ishiguro, M., Kitazawa, K., Takata, R., Ichinose, H., Kaneko, S., Igarashi, K., Samejima, M., & Tsumuraya, Y. (2011) J. Biol. Chem., 286, 27848–27854.
19) Takata, R., Tokita, K., Mori, S., Shimoda, R., Harada, N., Ichinose, H., Kaneko, S., Igarashi, K., Samejima, M., Tsumuraya, Y., & Kotake, T. (2010) Carbohydr. Res., 345, 2516–2522.
20) Gamauf, C., Marchetti, M., Kallio, J., Puranen, T., Vehmaanperä, J., Allmaier, G., Kubicek, C.P., & Seiboth, B. (2007) FEBS J., 274, 1691–1700.
21) Sekimata, M., Ogura, K., Tsumuraya, Y., Hashimoto, Y., & Yamamoto, S. (1989) Plant Physiol., 90, 567–574.
22) Uesaka, E., Sato, M., Raiju, M., & Kaji, A. (1978) J. Bacteriol., 133, 1073–1077.
23) Hata, K., Tanaka, M., Tsumuraya, Y., & Hashimoto, Y. (1992) Plant Physiol., 100, 388–396.
24) Ichinose, H., Fujimoto, Z., Honda, M., Harazono, K., Nishimoto, Y., Uzura, A., & Kaneko, S. (2009) J. Biol. Chem., 284, 25097–25106.
25) Ichinose, H., Fujimoto, Z., & Kaneko, S. (2013) Biosci. Biotechnol. Biochem., 77, 213–216.
26) Wu, Y., Williams, M., Bernard, S., Driouich, A., Showalter, A.M., & Faik, A. (2010) J. Biol. Chem., 285, 13638–13645.
27) Konishi, T., Kotake, T., Soraya, D., Matsuoka, K., Koyama, T., Kaneko, S., Igarashi, K., Samejima, M., & Tsumuraya, Y. (2008) Carbohydr. Res., 343, 1191–1201.
28) Eudes, A., Mouille, G., Thévenin, J., Goyallon, A., Minic, Z., & Jouanin, L. (2008) Plant Cell Physiol., 49, 1331–1341.
29) Tsumuraya, Y., Mochizuki, N., Hashimoto, Y., & Kováč, P. (1990) J. Biol. Chem., 265, 7207–7215.
30) Kotake, T., Dina, S., Konishi, T., Kaneko, S., Igarashi, K., Samejima, M., Watanabe, Y., Kimura, K., & Tsumuraya, Y. (2005) Plant Physiol., 138, 1563–1576.
31) Yoshimi, Y., Sugawara, Y., Hori, C., Igarashi, K., Kaneko, S., Tsumuraya, Y., & Kotake, T. (2017) Biosci. Biotechnol. Biochem., 81, 475–481.
32) Reiter, W.-D. (2008) Curr. Opin. Plant Biol., 11, 236–243.
33) Konishi, T., Takeda, T., Miyazaki, Y., Ohnishi-Kameyama, M., Hayashi, T., O’Neill, M.A., & Ishii, T. (2007) Glycobiology, 17, 345–354.
34) Kieliszewski, M.J. & Lamport, D.T.A. (1994) Plant J., 5, 157–172.
35) Ogawa-Ohnishi, M. & Matsubayashi, Y. (2015) Plant J., 81, 736–746.
36) Basu, D., Wang, W., Ma, S., DeBrosse, T., Poirier, E., Emch, K., Soukup, E., Tian, L., & Showalter, A.M. (2015) PLoS ONE, 10, e0125624.
37) Qu, Y., Egelund, J., Gilson, P.R., Houghton, F., Gleeson, P.A., Schultz, C.J., & Bacic, A. (2008) Plant Mol. Biol., 68, 43–59.
38) Dilokpimol, A., Poulsen, C.P., Vereb, G., Kaneko, S., Schulz, A., & Geshi, N. (2014) BMC Plant Biol., 14, 90.
39) Geshi, N., Johansen, J.N., Dilokpimol, A., Rolland, A., Belcram, K., Verger, S., Kotake, T., Tsumuraya, Y., Kaneko, S., Tryfona, T., Dupree, P., Scheller, H.V., Höfte, H., & Mouille, G. (2013) Plant J., 76, 128–137.
40) Knoch, E., Dilokpimol, A., Tryfona, T., Poulsen, C.P., Xiong, G., Harholt, J., Petersen, B.L., Ulvskov, P., Hadi, M.Z., Kotake, T., Tsumuraya, Y., Pauly, M., Dupree, P., & Geshi, N. (2013) Plant J., 76, 1016–1029.
41) Endo, M., Kotake, T., Watanabe, Y., Kimura, K., & Tsumuraya, Y. (2013) Planta, 238, 1157–1169.
42) Tryfona, T., Theys, T.E., Wagner, T., Stott, K., Keegstra, K., & Dupree, P. (2014) PLoS ONE, 9, e93291.
43) Liang, Y., Basu, D., Pattathil, S., Xu, W.L., Venetos, A., Martin, S.L., Faik, A., Hahn, M.G., & Showalter, A.M. (2013) J. Exp. Bot., 64, 5537–5551.
44) Cheung, A.Y., Wang, H., & Wu, H.M. (1995) Cell, 82, 383–393.
45) Wu, H.M., Wang, H., & Cheung, A.Y. (1995) Cell, 82, 395–403.
46) Mizukami, A.G., Inatsugi, R., Jiao, J., Kotake, T., Kuwata, K., Ootani, K., Okuda, S., Sankaranarayanan, S., Sato, Y., Maruyama, D., Iwai, H., Garénaux, E., Sato, C., Kitajima, K., Tsumuraya, Y., Mori, H., Yamaguchi, J., Itami, K., Sasaki, N., & Higashiyama, T. (2016) Curr. Biol., 26, 1091–1097.
47) Okuda, S., Tsutsui, H., Shiina, K., Sprunck, S., Takeuchi, H., Yui, R., Kasahara, R.D., Hamamura, Y., Mizukami, A., Susaki, D., Kawano, N., Sakakibara, T., Namiki, S., Itoh, K., Otsuka, K., Matsuzaki, M., Nozaki, H., Kuroiwa, T., Nakano, A., Kanaoka, M.M., Dresselhaus, T., Sasaki, N., & Higashiyama, T. (2009) Nature, 458, 357–361.
48) Motose, H., Sugiyama, M., & Fukuda, H. (2004) Nature, 429, 873–878.
49) Shi, H., Kim, Y., Guo, Y., Stevenson, B., & Zhu, J.K. (2003) Plant Cell, 15, 19–32.
50) Sardar, H.S., Yang, J., & Showalter, A.M. (2006) Plant Physiol., 142, 1469–1479.
51) Nguema-Ona, E., Bannigan, A., Chevalier, L., Baskin, T.I., & Driouich, A. (2007) Plant J., 52, 240–251.
著者紹介Author Profile
 円谷 陽一(つむらや よういち)
円谷 陽一(つむらや よういち)埼玉大学名誉教授.学術博士.
略歴1950年福島県に生れる.73年埼玉大学理工学部生化学科卒業.東京農工大学大学院修士課程,大阪市立大学大学院博士課程等を経て,80年埼玉大学理学部生化学科助手.理工学研究科生命科学部門教授を経て2016年定年退職.
研究テーマと抱負大学院生,デンプン会社勤務,埼玉大学勤務,海外留学時とも主に微生物・植物の糖質生化学・糖鎖生物学に興味を抱いて研究に取り組んだ.長い期間,研究活動を楽しめたと感じている.
趣味囲碁,写真撮影.
 小竹 敬久(こたけ としひさ)
小竹 敬久(こたけ としひさ)埼玉大学大学院理工学研究科教授.博士(学術).
略歴1972年岐阜県に生る.2000年3月広島大学大学院生物圏科学研究科修了・同年4月岡山県生物科学総合研究所流動研究員.01年7月埼玉大学理学部助手,09年1月同大学院理工学研究科准教授,17年4月より現職.
研究テーマと抱負植物の糖鎖生物学.植物の生命現象を糖鎖の働きから解明したい.
ウェブサイトhttp://park.saitama-u.ac.jp/~seitaibusshitsu/index.html
趣味散歩,自然観察,スポーツ全般.