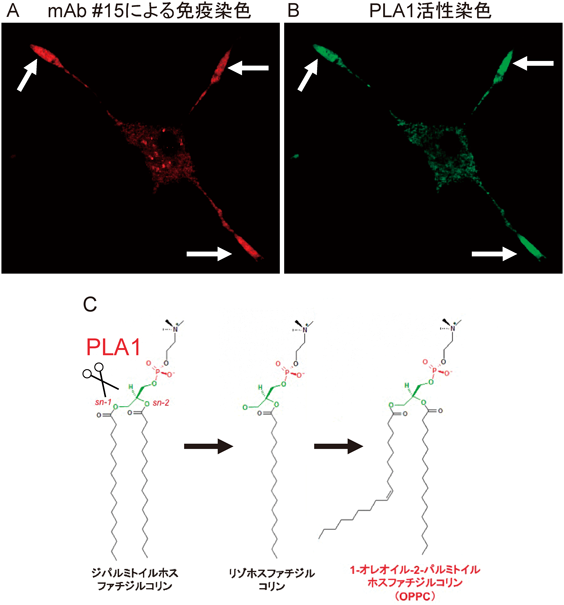
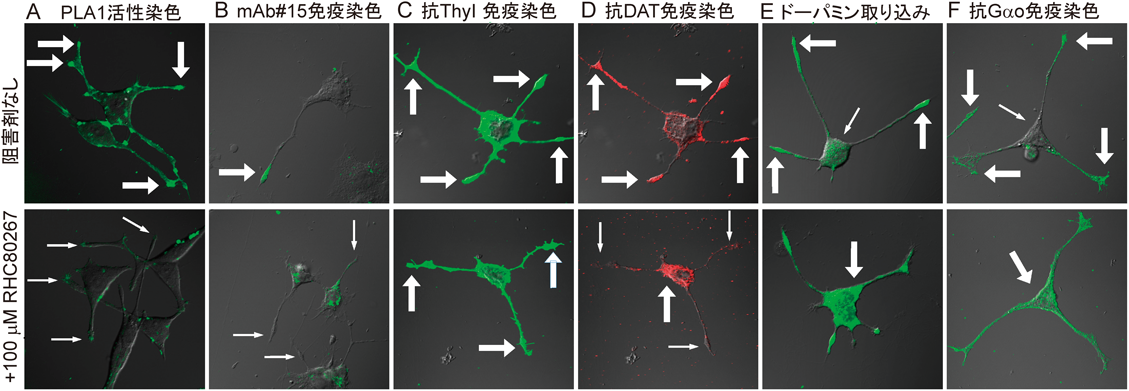
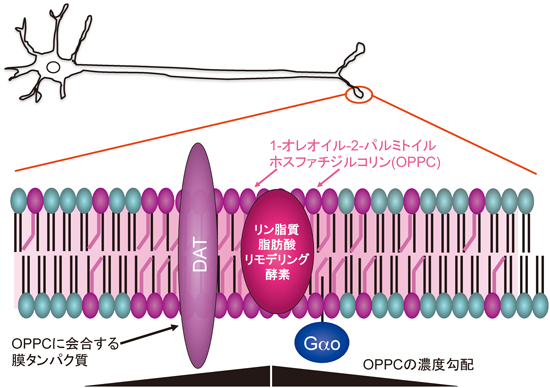
sn-1位に不飽和脂肪酸をもつリン脂質分子種がつくる神経細胞膜機能ドメインMembrane domains in a neuron marked by rare phospholipids with unsaturated fatty acid at the sn-1 site
高知大学医学部生化学教室Department of Biochemistry, Kochi University Medical School ◇ 高知県南国市岡豊町小蓮 ◇ Kohasu, Okocyou, Nankoku, Kochi 783–8505, Japan
発行日:2017年8月25日Published: August 25, 2017