細胞の内部は,水よりもむしろタンパク質や核酸などにより埋めつくされ混雑している.この状態は分子混雑と呼ばれ,細胞の状態を決める重要な要素の一つと考えられている1).たとえば,細胞内のタンパク質濃度は,約300~350 mg/mLと見積もられているが,このような高タンパク質濃度は,タンパク質の構造の不安定化を引き起こしたり,逆に安定化したり,酵素活性など機能に影響を与える.つまり,生化学実験等の理想環境下における試験管実験系と実際の細胞内とには大きな乖離がある.両者をつなげるためには,細胞内の分子混雑を知ることが必要不可欠と考えられており,古くから分子混雑のタンパク質への影響は研究されている.
これまで細胞内の分子混雑状態は,FRAP(fluorescence recovery after photobleaching)法や,FCS(fluorescence correlation spectroscopy)法など,蛍光タンパク質や色素などのプローブの流動性を指標として評価されてきた2).しかしながら,細胞内には細胞骨格やクロマチンのようなタンパク質が集まった構造体が存在し,この流動性に影響を与える可能性がある.細胞内分子混雑を評価するためには,プローブの動き(流動性)のみでは不十分であり,他の要素,たとえば,分子・タンパク質密度などの情報(混雑具合)が必要である.この問題を解決すべく,我々は,周囲のタンパク質や核酸濃度に依存して蛍光波長が変化する蛍光タンパク質GimRETを開発し,生きた細胞内で分子の混雑具合と流動性を同時計測することに成功した3).本稿では,GimRETの開発戦略とGimRETを用いた細胞内分子混雑計測の実例を記す.
1)蛍光タンパク質の疎水性感受性を高める
GimRETは,蛍光波長が異なる二つの蛍光タンパク質から構成され,溶液内の分子混雑具合に依存して,その蛍光スペクトルが変化する環境感受性蛍光プローブである.有機色素を基盤とした分子混雑感受性色素も存在するが,細胞や個体などへの導入には困難がある.一方,蛍光タンパク質を基盤としたプローブは,遺伝子にコードした形で細胞内に導入できるため,組織内や個体内の細胞でも,観察試料を生かしたままその蛍光を検出できる.上記が,我々が蛍光タンパク質をGimRETの基盤とした理由である.
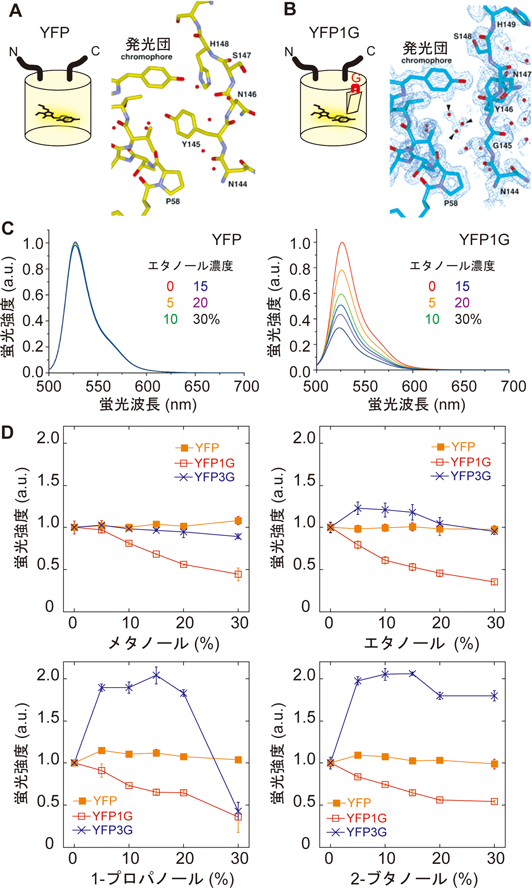
蛍光タンパク質は,2008年ノーベル化学賞受賞者である下村脩博士により1960年代にオワンクラゲから発見された4).後に,遺伝子工学技術により蛍光強度や波長特性が異なる改変蛍光タンパク質が作られ,また,サンゴ等他の生物からも発見され,現在,さまざまな種類の蛍光タンパク質が存在する5, 6).蛍光タンパク質の構造は,由来によらずほぼ同様の円柱構造をしており,蛍光を発する発光団はその円柱の中心にある(図1A,左).円柱構造により,発光団と水分子との相互作用が制限されているため,発光団は溶液中の環境変化に対し比較的安定した蛍光を発することができる.逆にいえば,円柱構造に変異を与えることで,蛍光を不安定化させることができる.
我々はここで,タンパク質濃度が溶液の疎水性と直接関与していることに着目した.溶液の疎水性が,分子混雑具合を評価する上で最大のファクターであるタンパク質濃度とみなせると考えたのである.実際に疎水性感受性の有機蛍光色素は,溶液内のタンパク質濃度により,その蛍光波長・強度が変化する3).一方で,通常の蛍光タンパク質の蛍光は,円柱構造の防御機構により,溶液の疎水性に対して安定している.第一の開発は,蛍光タンパク質に,疎水性感受性を持たせることであった.
我々のアイデアは,蛍光タンパク質の円柱構造に「小さな穴」をあけ,蛍光が失われない程度に発光団と水分子を相互作用させることであった.蛍光タンパク質の分子構造をよくながめてみると,発光団の近くに145番目チロシン残基(用いたオワンクラゲ由来の蛍光タンパク質は全238残基)のフェノール環が配置されており,これが水分子の発光団への接近を妨げている(図1A,右).我々は,144番目と145番目のアミノ酸残基の間に,新たにアミノ酸を挿入し,チロシン残置を少しだけ移動させることを試みた(図1B,左).3種類の色の異なる蛍光タンパク質(青,緑,黄)にさまざまなアミノ酸残基を挿入してみたが,ここでは,最終的に使用した黄色蛍光タンパク質(YFP)にグリシンを挿入した場合(YFP1G)のみを記す.
我々は,グリシンが挿入されたYFPを構築した後,大阪大学今田勝巳教授にその結晶構造解析を依頼した.結晶構造解析によれば,我々の期待どおりに,グリシン挿入は確かに145番目チロシン残基の向きを変え,発光団の近くには水分子が配置されていた(図1B,右).YFPおよびYFP1Gの蛍光のエタノール濃度依存性を調べてみると,YFPの蛍光はエタノール濃度に依存することなく安定した蛍光を発していたが,YFP1Gの蛍光はエタノール濃度の増加に伴い蛍光強度が低下していた(図1C).さまざまな有機溶媒に対しての依存性を調べると,どの有機溶媒に対してもYFPの蛍光は変化せず(図1D,黄),一方,YFP1Gは濃度に対してほぼ線形的な減少を見せた(図1D,赤).面白いことに,グリシンを三つ挿入したYFP(YFP3G)の蛍光強度は,有機溶媒の種類により異なる反応を見せる,二相性の反応が確認される(図1D,青)など,疎水性以外の要素にも影響を受けていた.本開発の「キモ」は,グリシン残基を「一つだけ」挿入したことであった.
2)蛍光共鳴エネルギー移動法を適応する
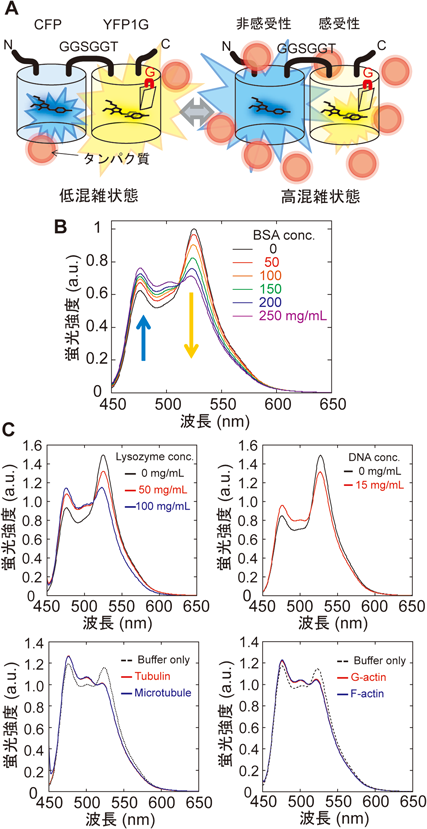
蛍光強度は,計測する指標としては使いにくい.なぜなら,蛍光タンパク質の細胞内の発現量は,細胞によって異なるからである.我々は,分子混雑の変化を蛍光強度変化ではなく蛍光スペクトル変化として検出できるようにするために,蛍光共鳴エネルギー移動法(Förster resonance energy transfer:FRET)を適用した7, 8).YFP1Gに疎水性感受性を持たない青色蛍光タンパク質(CFP)を短いリンカーペプチドで融合させる(図2A).CFPとYFP1Gがごく近傍にある場合に,CFPが吸収した光エネルギーの一部は蛍光を発することなくYFP1Gへと移動し,YFP1Gが代わりに蛍光を発する.YFP1Gは溶液内の疎水性に感受する一方でCFPは感受しないので,この融合蛍光タンパク質は,疎水性が上昇すると黄色蛍光が減少し青色蛍光が増加する.すなわち,CFP–YFP1G融合タンパク質は,溶液の疎水性に依存して蛍光スペクトルが変化するプローブとなる.我々は,この新しい蛍光タンパク質を“GimRET(glycine inserted mutant FRET sensor)”と名づけた.
GimRETは,確かにウシ血清アルブミン(bovine serum albumin:BSA)濃度に依存して,その蛍光スペクトルの形状を変化させた(図2B).GimRETはBSAのみならず,リゾチームやアクチン,チュブリン,さらには,核酸にも反応を見せた.面白いことに,アクチンやチュブリンの重合化には反応しないことから,GimRETは,タンパク質の超構造にはあまり反応せず濃度に対して優位に反応していると考えられる(図2C).また,溶液粘性を調べる際の標準分子とされるポリエチレングリコールやスクロース等の小分子には反応しなかった3).GimRETは,高分子(macromolecule)の混雑具合に反応するプローブであるといえる.
3)実証実験
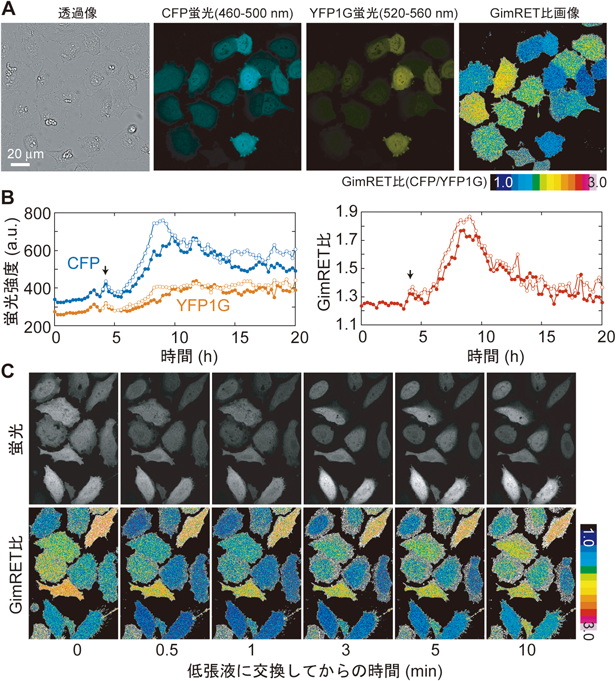
試験管実験により,GimRETの蛍光スペクトルのタンパク質濃度感受性が示された.次は,細胞内部のタンパク質濃度すなわち混雑具合の変化をGimRETで検出できることを示さなければならない.我々は,GimRETをヒト子宮頸がん由来のHeLa細胞株に発現させ培養し,そのようすを共焦点蛍光顕微鏡で観察した.GimRETは,周囲の混雑具合に依存して,CFPが発する蛍光とYFP1Gが発する蛍光が逆行的に変化する(図2B, C).すなわち,レーザー光照射によりCFPを励起し,波長460~500 nmおよび波長520~560 nmの二つの蛍光画像を取得し,その強度比(CFP/YFP1G,以下GimRET比と書く)を計算することで,細胞内の混雑具合を捉えることができる(図3A).図3A左は,GimRET比の値に疑似色(黒–青–緑–黄–赤–白)を対応させた画像であり,この図における色はGimRETの発光色ではないことに注意されたい.細胞は分裂直後から次の分裂時のためにタンパク質を合成し,容積を増やした後に次の分裂を開始する.実際にGimRET比をみてみると,細胞分裂直後からGimRET比は増加し,その後,ゆっくりと減少している(図3B).薬剤添加によりタンパク質合成あるいは分解を阻害すると,GimRET比はそれに準じた変化を見せることも確認できている3).以上の結果から,GimRETは細胞内のタンパク質濃度の変化によって,その蛍光波長を変化させていると考えられる.
しかし,これだけでは実証にはならない.我々は,さらに,細胞の容積を強制的に変化させたときのタンパク質濃度変化の検出を試みた.具体的には,細胞培養液を高張液から低張液に置換し,浸透圧変化による細胞膨張により,強制的に細胞内のタンパク質濃度を変化させた.実際に,溶液交換後すぐに,GimRET比が低下し,その後に徐々に回復しているようすが確認できた(図3C).逆に,細胞骨格を破壊し,強制的に細胞の容積を圧縮させると,GimRET比が増加することも確認してある3).このように,細胞内においてもGimRETは,混雑具合の変化に対して感受性があることが示された.
1)GimRETによる分子混雑計測と拡散計測との比較
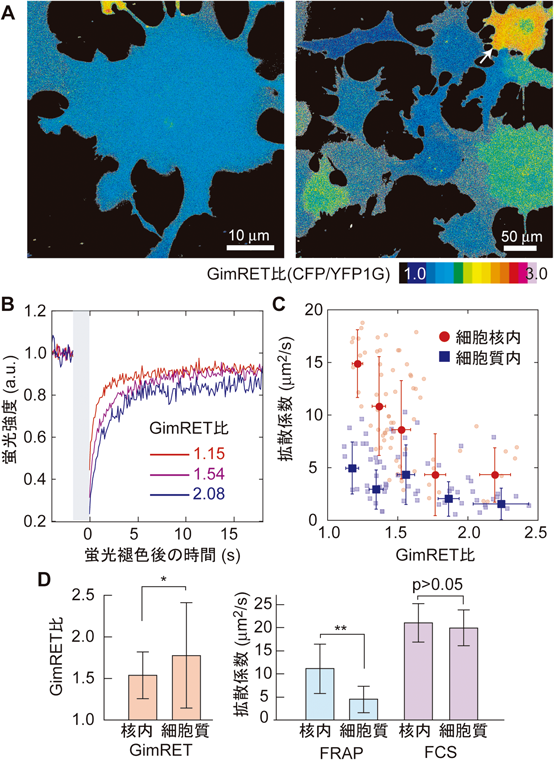
我々は,GimRETを用いて,生きた細胞内で混雑具合と流動性を同時に計測した.具体的には,GimRET比により細胞内の混雑具合を,GimRETの拡散速度によりその流動性を計測した.アフリカミドリザル腎臓由来のCOS-7細胞にGimRETを発現させ,上述と同様に共焦点蛍光顕微鏡を用いたが,今回はより高倍率の対物レンズを用いて観察した.ほとんどの細胞において細胞核内と細胞質内とでGimRET比に有意な差はみられず,分子混雑状態は細胞内でほぼ一様であると考えられる(図4A).まれに,細胞質内のGimRET比が高い細胞がみられるため,細胞質の方が細胞核内に比べ平均的に少し分子混雑が高いといえる(図4Aの矢印,図4D,左).
一方で,GimRETは蛍光タンパク質であり,細胞内にて容易にFRAPが適用できる.FRAP法とは,強い光照射により細胞の一部のみ蛍光を退色させ,その後の蛍光強度回復の速度により,蛍光タンパク質の拡散の速度を算出する方法である9).細胞によって,GimRET比はばらつきを見せ,蛍光強度の回復速度すなわちGimRETの拡散速度は,確かにGimRET比と相関があった(図4B).面白いことに,この相関は,細胞核内と細胞質内で異なっていた(図4C).正しくは,細胞核内と細胞質内では,FRAPにより算出された拡散係数が異なっていた.GimRETが感受できるのは,GimRET自身の近傍の分子混雑具合のみであり,一方で,FRAP計測はより広範囲にわたる拡散を捉えている.そこで,我々は,北海道大学金城政孝教授にFCSによるGimRETの拡散速度計測をお願いした.FCSは,レーザー集光点のみにおける計測すなわち狭範囲での計測となる10).FCSにより算出された拡散係数は,FRAPで産出された値より高く,また,細胞核内と細胞質内で大きな差はみられなかった(図4D).これらの結果を総合的に解釈すると,細胞質内では比較的大きな構造体によりタンパク質の拡散が抑制されており,細胞核内は比較的自由な拡散が許されている環境である,といえる.このようにGimRETは,細胞内における分子混雑の混雑具合と流動性とを分けて議論することを可能にする.
2)細胞分化過程における細胞核内の分子混雑変化
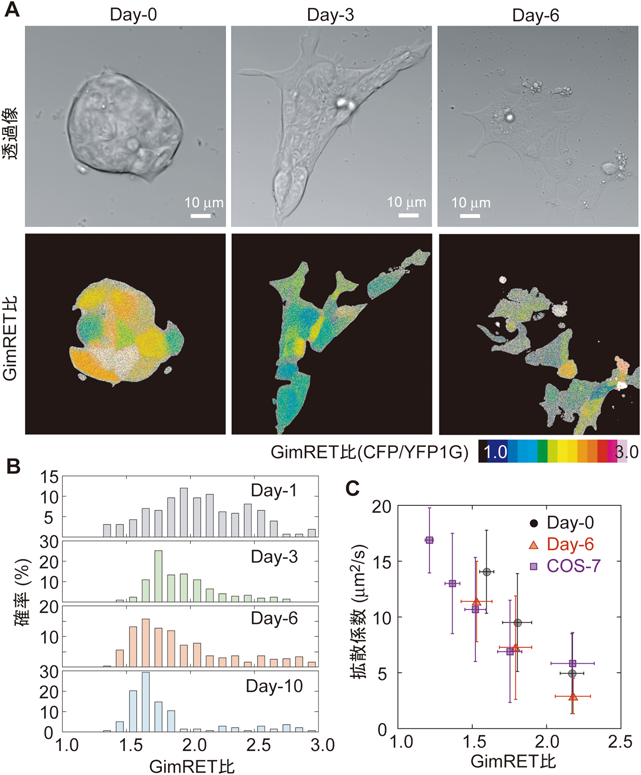
細胞分化は,細胞核内部の状況が大きく変化する現象の一つである.我々は,マウスの胚性幹細胞(ES細胞)の初期分化過程における,細胞核内の分子混雑変化をGimRETで評価してみた(図5A).分化誘導前のES細胞は,高い混雑具合を示し,また,細胞間のばらつきも大きかった(図5B,最上段).分化誘導後は,日が経つにつれ,混雑具合は減少し,そのばらつきも小さくなっていった(図5B).一方で,FRAPによる拡散速度計測では,分化誘導前のES細胞の核内は分化誘導後に比べ拡散係数が高い値を示し,混雑具合(GimRET比)とは逆の結果となった(図5C).分化前のES細胞の細胞核内は分子が多く混雑している一方で,その中の流動性は高く保たれていたのである.これは,クロマチンの構造とその中でのタンパク質の流動性との関係を表しているのかもしれない.未分化時には,核内のクロマチンはオープン構造(ほどけた構造)をしている11).GimRETは局所の分子混雑しか検出できないので,クロマチンの凝集に伴いGimRET比は低下していく.一方で,ほどけたクロマチン構造は,分子の自由拡散を保障していることが知られており,クロマチンの凝集に伴い拡散係数は低下する12).このように,分子混雑状態と流動性とを分けて計測することにより,細胞核内部のようすをより詳細にイメージすることができる.
3)GimRET融合タンパク質による選択的分子混雑観察
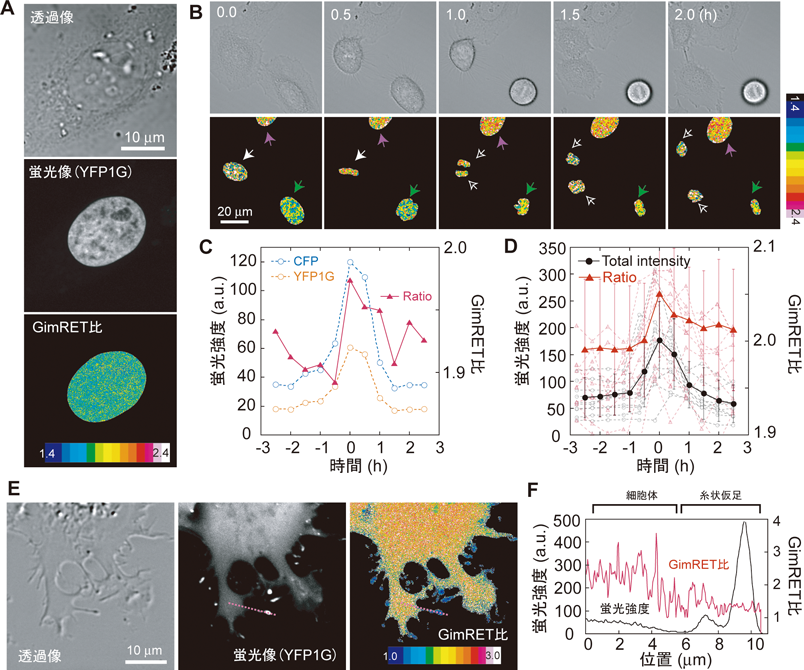
GimRETは蛍光タンパク質であるため,通常の蛍光タンパク質同様に,観察したい標的タンパク質に融合させることで,その標的タンパク質近傍の分子混雑状態を選択的に観察することができる.我々は,ヒストンタンパク質の一つであるH2BにGimRETを融合させ,細胞分裂中の染色体凝集時におけるヒストン周辺の分子混雑を観察してみた.細胞核内においてH2B周辺の混雑具合は一様であった(図6A).細胞分裂に伴い,染色体は凝集,分裂,逆凝集と,ダイナミックな形態変化を見せる(図6B).染色体凝集時に,わずかではあるが確実に,H2B–GimRET融合体のGimRET比の変化が観察された(図6C).数サンプルのGimRET比の変化を収集し平均化することにより,染色体凝集前後におけるGimRET比増減のようすが確認できる(図6D).H2Bの凝集具合は蛍光強度変化により評価でき(図6D,黒),染色体凝集後は凝集前に比べて,H2Bの周辺は混雑していると思われる(図6D,赤).
他の例として,モータータンパク質の一つであるミオシンXにGimRETを融合し,観察してみた(図6E).ミオシンXは,仮足伸長に必須のタンパク質であり,細胞質内から糸状仮足先端に向かい移動する13).面白いことに,ミオシンXは,糸状仮足内部のアクチンフィラメントにのみ選択的に結合することが知られている.ミオシンX−GimRET融合体のGimRET比を観察してみると,細胞質内(正しくは葉状仮足内)に比べ糸状仮足内において,低いGimRET比値が確認された(図6F).ミオシンX周辺の分子混雑は,糸状仮足内の方が低く,タンパク質が輸送されるために障害が少ないのかもしれない.
上述した二つの実験例は,まだまだ始めたばかりであるが,GimRET融合タンパク質が,その局在とともに周辺局部の分子混雑の情報を与えてくれることを示しており,GimRETの今後のさまざまな研究分野への応用展開が期待される.
このように,我々は,非常に単純な遺伝子改変により,これまでにない蛍光タンパク質プローブを開発し,これまで直接的に測ることができなかった細胞内の分子混雑を,蛍光顕微鏡で容易に評価できることを示した.
しかしながら,問題は残っている.YFPに施したグリシン挿入が発光団に疎水感受性をもたらしたメカニズムが解けていないままなのである.そもそも分子混雑がタンパク質の構造折りたたみや機能に与える影響などにはまだまだ不明な点が多く,今後の研究を待たなければならない.GimRETとは異なる戦略で,CFPとYFPの間に,分子混雑により構造が変化すると予想されるペプチドを挿入した分子混雑プローブが他グループにより開発されているが14),そのメカニズムにもまだ不明な点が残っている.GimRETは,プローブとして有効であるとともに,生化学分野においても,蛍光タンパク質の発光メカニズムの解明という観点で,よい研究材料かもしれない.
引用文献References
1) Goodsell, D.S.(2009) The Machinery of Life, 2nd ed., Springer, New York.
2) van den Berg, B., Ellis, R.J., & Dobson, C.M. (1999) EMBO J., 18, 6927–6933.
3) Morikawa, T.J., Fujita, H., Kitamura, A., Horio, T., Yamamoto, J., Kinjo, M., Sasaki, A., Machiyama, H., Yoshizawa, K., Ichimura, T., Imada, K., Nagai, T., & Watanabe, T.M. (2016) Sci. Rep., 6, 22342.
4) Shimomura, O., Johnson, F.H., & Saiga, Y. (1962) J. Cell. Comp. Physiol., 59, 223–239.
5) Tsien, R.Y. (1998) Annu. Rev. Biochem., 67, 509–544.
6) Chudakov, D.M., Matz, M.V., Lukyanov, S., & Lukyanov, K.A. (2010) Physiol. Rev., 90, 1103–1163.
7) Förster, T. (1948) Ann. Phys., 437, 55–57.
8) Felber, L.M., Cloutier, S.M., Kündig, C., Kishi, T., Brossard, V., Jichlinski, P., Leisinger, H.J., & Deperthes, D. (2004) Biotechnique, 36, 878–885.
9) Axelrod, D., Koppel, D., Schlessinger, J., Elson, E., & Webb, W.W. (1976) Biophys. J., 16, 1055–1069.
10) Magde, D., Elson, E.L., & Webb, W.W. (1974) Biopolymers, 13, 29–61.
11) Gaspar-Maia, A., Alajem, A., Meshorer, E., & Ramalho-Santos, M. (2011) Nat. Rev. Mol. Cell Biol., 12, 36–47.
12) Bancaud, A., Huet, S., Daigle, N., Mozziconacci, J., Beaudouin, J., & Ellenberg, J. (2009) EMBO J., 28, 3785–3798.
13) Berg, J.S. & Cheney, R.E. (2002) Nat. Cell Biol., 4, 246–250.
14) Boersma, A.J., Zuhorn, I.S., & Poolman, B.A. (2015) Nat. Methods, 12, 227–229.