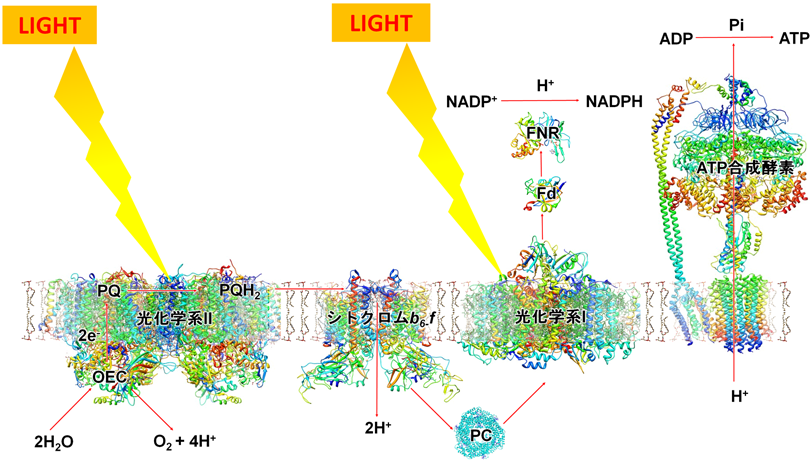
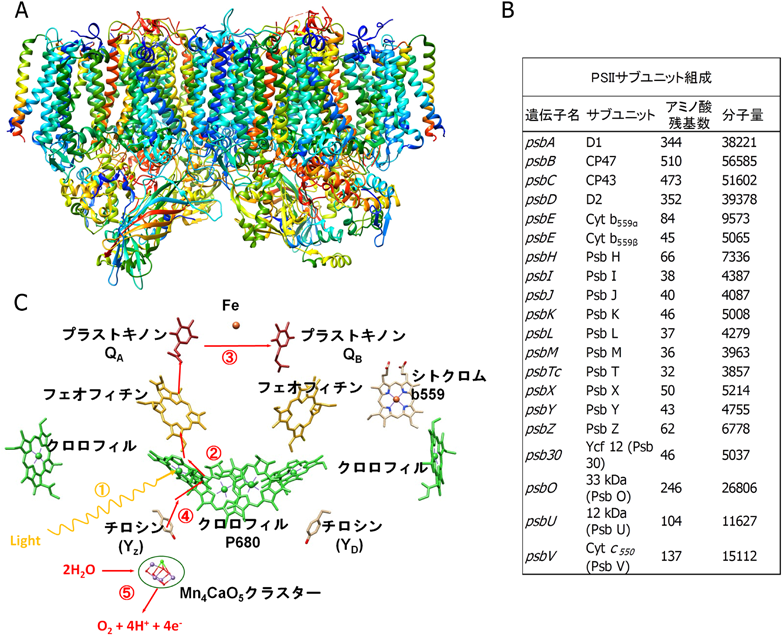
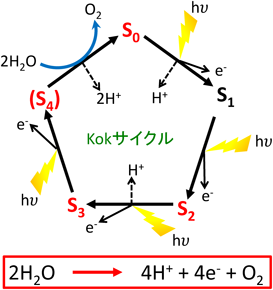
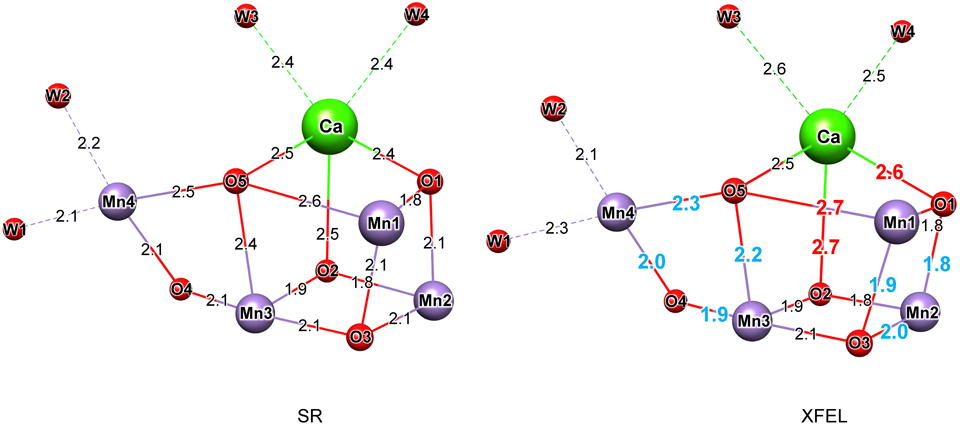
11) Yano, J., Kern, J., Sauer, K., Latimer, M.J., Pushkar, Y., Biesiadka, J., Loll, B., Saenger, W., Messinger, J., Zouni, A., & Yachandra, V.K. (2006) Science, 314, 821–825.
17) Kern, J., Alonso-Mori, R., Hellmich, J., Tran, R., Hattne, J., Laksmono, H., Glockner, C., Echols, N., Sierra, R.G., Sellberg, J., Lassalle-Kaiser, B., Gildea, R.J., Glatzel, P., Grosse-Kunstleve, R.W., Latimer, M.J., McQueen, T.A., DiFiore, D., Fry, A.R., Messerschmidt, M., Miahnahri, A., Schafer, D.W., Seibert, M.M., Sokaras, D., Weng, T.C., Zwart, P.H., White, W.E., Adams, P.D., Bogan, M.J., Boutet, S., Williams, G.J., Messinger, J., Sauter, N.K., Zouni, A., Bergmann, U., Yano, J., & Yachandra, V.K. (2012) Proc. Natl. Acad. Sci. USA, 109, 9721–9726.
18) Kern, J., Alonso-Mori, R., Tran, R., Hattne, J., Gildea, R.J., Echols, N., Glockner, C., Hellmich, J., Laksmono, H., Sierra, R.G., Lassalle-Kaiser, B., Koroidov, S., Lampe, A., Han, G., Gul, S., Difiore, D., Milathianaki, D., Fry, A.R., Miahnahri, A., Schafer, D.W., Messerschmidt, M., Seibert, M.M., Koglin, J.E., Sokaras, D., Weng, T.C., Sellberg, J., Latimer, M.J., Grosse-Kunstleve, R.W., Zwart, P.H., White, W.E., Glatzel, P., Adams, P.D., Bogan, M.J., Williams, G.J., Boutet, S., Messinger, J., Zouni, A., Sauter, N.K., Yachandra, V.K., Bergmann, U., & Yano, J. (2013) Science, 340, 491–495.
19) Kern, J., Tran, R., Alonso-Mori, R., Koroidov, S., Echols, N., Hattne, J., Ibrahim, M., Gul, S., Laksmono, H., Sierra, R.G., Gildea, R.J., Han, G., Hellmich, J., Lassalle-Kaiser, B., Chatterjee, R., Brewster, A.S., Stan, C.A., Glockner, C., Lampe, A., DiFiore, D., Milathianaki, D., Fry, A.R., Seibert, M.M., Koglin, J.E., Gallo, E., Uhlig, J., Sokaras, D., Weng, T.C., Zwart, P.H., Skinner, D.E., Bogan, M.J., Messerschmidt, M., Glatzel, P., Williams, G.J., Boutet, S., Adams, P.D., Zouni, A., Messinger, J., Sauter, N.K., Bergmann, U., Yano, J., & Yachandra, V.K. (2014) Nat. Commun., 5, 4371.
20) Kupitz, C., Basu, S., Grotjohann, I., Fromme, R., Zatsepin, N.A., Rendek, K.N., Hunter, M.S., Shoeman, R.L., White, T.A., Wang, D., James, D., Yang, J.-H., Cobb, D.E., Reeder, B., Sierra, R.G., Liu, H., Barty, A., Aquila, A.L., Deponte, D., Kirian, R.A., Bari, S., Bergkamp, J.J., Beyerlein, K.R., Bogan, M.J., Caleman, C., Chao, T.-C., Conrad, C.E., Davis, K.M., Fleckenstein, H., Galli, L., Hau-Riege, S.P., Kassemeyer, S., Laksmono, H., Liang, M., Lomb, L., Marchesini, S., Martin, A.V., Messerschmidt, M., Milathianaki, D., Nass, K., Ros, A., Roy-Chowdhury, S., Schmidt, K., Seibert, M., Steinbrener, J., Stellato, F., Yan, L., Yoon, C., Moore, T.A., Moore, A.L., Pushkar, Y., Williams, G.J., Boutet, S., Doak, R.B., Weierstall, U., Frank, M., Chapman, H.N., Spence, J.C.H., & Fromme, P. (2014) Nature, 513, 261–265.
21) Sauter, N.K., Echols, N., Adams, P.D., Zwart, P.H., Kern, J., Brewster, A.S., Koroidov, S., Alonso-Mori, R., Zouni, A., Messinger, J., Bergmann, U., Yano, J., & Yachandra, V.K. (2016) Nature, 533, E1–E2.
22) Hirata, K., Shinzawa-Itoh, K., Yano, N., Takemura, S., Kato, K., Hatanaka, M., Muramoto, K., Kawahara, T., Tsukihara, T., Yamashita, E., Tono, K., Ueno, G., Hikima, T., Murakami, H., Inubushi, Y., Yabashi, M., Ishikawa, T., Yamamoto, M., Ogura, T., Sugimoto, H., Shen, J.R., Yoshikawa, S., & Ago, H. (2014) Nat. Methods, 11, 734–746.
23) Suga, M., Akita, F., Hirata, K., Ueno, G., Murakami, H., Nakajima, Y., Shimizu, T., Yamashita, K., Yamamoto, M., Ago, H., & Shen, J.-R. (2015) Nature, 517, 99–103.
25) Yamanaka, S., Isobe, H., Kanda, K., Saito, T., Umena, Y., Kawakami, K., Shen, J.-R., Kamiya, N., Okumura, M., Nakamura, H., & Yamaguchi, K. (2011) Chem. Phys. Lett., 511, 138–145.
32) Rapatskiy, L., Cox, N., Savitsky, A., Ames, W.M., Sander, J., Nowaczyk, M.M., Rögner, M., Boussac, A., Neese, F., Messinger, J., & Lubitz, W. (2012) J. Am. Chem. Soc., 134, 16619–16634.
33) Cox, N., Rapatskiy, L., Su, J.H., Pantazis, D.A., Sugiura, M., Kulik, L., Dorlet, P., Rutherford, A.W., Neese, F., Boussac, A., Lubitz, W., & Messinger, J. (2011) J. Am. Chem. Soc., 133, 3635–3648.
39) Najafpour, M.M., Renger, G., Holyńska, M., Moghaddam, A., Aro, E.-M., Carpentier, R., Nishihara, H., Eaton-Rye, J.J., Shen, J.-R., & Allakhverdiev, S.I. (2016) Chem. Rev., 116, 2886–2936.
40) Sugahara, M., Mizohata, E., Nango, E., Suzuki, M., Tanaka, T., Masuda, T., Tanaka, R., Shimamura, T., Tanaka, Y., Suno, C., Ihara, K., Pan, D., Kakinouchi, K., Sugiyama, S., Murata, M., Inoue, T., Tono, K., Song, C., Park, J., Kameshima, T., Hatsui, T., Joti, Y., Yabashi, M., & Iwata, S. (2015) Nat. Methods, 12, 61–63.
41) Tono, K., Nango, E., Sugahara, M., Song, C., Park, J., Tanaka, T., Tanaka, R., Joti, Y., Kameshima, T., Ono, S., Hatsui, T., Mizohata, E., Suzuki, M., Shimamura, T., Tanaka, Y., Iwata, S., & Yabashi, M. (2015) J. Synchr. Rad., 22, 532–537.
42) Kubo, M., Nango, E., Tono, K., Kimura, T., Owada, S., Song, C., Mafuné, F., Miyajima, K., Takeda, Y., Kohno, J., Miyauchi, N., Nakane, T., Tanaka, T., Nomura, T., Davidsson, J., Tanaka, R., Murata, M., Kameshima, T., Hatsui, T., Joti, Y., Neutze, R., Yabashi, M., & Iwata, S. (2017) J. Synchr. Rad., 24, 1086–1091.
43) Suga, M., Akita, F., Sugahara, M., Kubo, M., Nakajima, Y., Nakane, T., Yamashita, K., Umena, Y., Nakabayashi, M., Yamane, T., Nakano, T., Suzuki, M., Masuda, T., Inoue, S., Kimura, T., Nomura, T., Yonekura, S., Yu, L.J., Sakamoto, T., Motomura, T., Chen, J.H., Kato, Y., Noguchi, T., Tono, K., Joti, Y., Kameshima, T., Hatsui, T., Nango, E., Tanaka, R., Naitow, H., Matsuura, Y., Yamashita, A., Yamamoto, M., Nureki, O., Yabashi, M., Ishikawa, T., Iwata, S., & Shen, J.R. (2017) Nature, 543, 131–135.