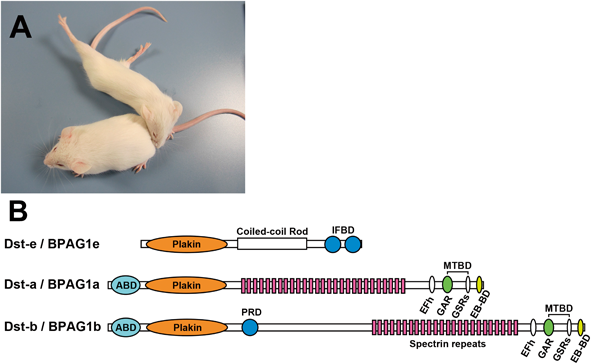
ジストニア様の運動症状と感覚神経変性を示すdystonia musculorumマウスの病態解析Neuropathological analysis of dystonia musculorum mice which exhibit motor disorder with dystonia-like movement and sensory neuropathy
新潟大学大学院医歯学総合研究科神経生物・解剖学Division of Neurobiology and Anatomy, Graduate School of Dental and Medical Sciences, Niigata University ◇ 〒951–8510 新潟市中央区旭町通1番町757 ◇ 1–757 Asahimachi, Chuo-ku, Niigata 951–8510, Japan
発行日:2017年10月25日Published: October 25, 2017