痛みの生化学 最近20年の進歩Biochemistry of pain, progress in these 20 years
関西医科大学医化学講座Department of Medical Chemistry, Kansai Medical University ◇ 〒573–1010 枚方市新町2–5–1 ◇ 2–5–1 Shin-machi, Hirakata, 573–1010
発行日:2017年12月25日Published: December 25, 2017
痛みはだれもが日常生活で常に体験するものであり,病院を訪れる患者の最も多い理由である.高齢社会を迎えた日本にあって,痛みは人々にとって最大の関心事であり,人類の克服すべき課題である.アリストテレスが「痛みは魂の苦悩」といったように,長い間,痛みは主観的,観念的であり,科学にはなりえないと考えられてきた.1997年にカプサイシン受容体が熱の侵害受容器として発見され,生化学的な研究がスタートしたのを契機として,1999年に痛みの分子機構について「痛みの生化学」と題して本誌に総説を執筆した(伊藤誠二,南敏明(1999)生化学,77, 17‒33).現在,痛みの感覚受容面だけでなく,情動認知面に関わる脳の痛みのネットワークが理解されつつある.本稿では,機能分子から動物個体にわたる痛みのメカニズムについて「痛みの生化学 最近20年の進歩」としてまとめる.
© 2017 公益社団法人日本生化学会© 2017 The Japanese Biochemical Society
「我思う,ゆえに我あり」と唱えた17世紀のフランスの哲学者デカルトは「観察できるヒトの行動は二つに分類できる:単純と複雑.単純な行動は,特定の感覚のように,いつも決まった同じパターンを引き起こす行動である.複雑な行動は対照的に感覚と行動が予期できず,意志の気まぐれに翻弄される行動である」と述べている.痛みは生物が持つ基本的な防衛反応の一つで,熱いものに触ったときは反射的に手を引っ込める単純な行動をとる.炎症や怪我をしたときには,痛みは増強,持続して,炎症部位や損傷部位の動きを抑えることにより,回復力を高め,治癒を促進する役割がある.一方,痛みは生殖や食事といったごく基本的な営みと同じく,種の保存,生命の維持に不可欠に機能である.兵士は戦場で傷を受けても痛みを感じる前に,敵と戦うか逃げるか判断する.このように,痛みには組織損傷に伴う痛みがどこで起こっているのかを識別する感覚受容の側面と,不安,恐怖,過去の記憶などの影響を受ける情動認知の側面の二面がある.
古代ギリシャの哲学者アリストテレスは感覚を視覚,聴覚,味覚,嗅覚と触覚の五つに分けたが,触覚は,触る,触られるだけでなく,温かさ,冷たさ,痒み,痛さなどの感覚を一くくりにした体性感覚のことである.さまざまな刺激に応答するために,体性感覚の受容器は全身すべての部位に分布し,外受容,内受容,固有知覚の三つに分類される.外受容は温刺激,冷刺激,触刺激,圧刺激など体の外から受ける感覚で,体に危害や障害を与える刺激で生じる痛覚も含む.内受容は内臓器官の機能や体の内部状態に関係する感覚で,その多くは意識されることはない.運動したときの血中の酸素濃度やpHの変化を内受容器が感知して,発汗,脈拍,血圧や呼吸などが自律神経系によってコントロールされる.固有知覚は自分自身の四肢や頭部の位置,姿勢,筋肉の動きに関係する感覚である.
皮膚,筋肉や内臓からの情報を集める感覚神経を出す神経細胞(ニューロン)は脊柱の後ろ側の出口付近にある後根神経節に存在する(図1).ニューロンからでた神経線維はすぐに枝分かれして皮膚や筋肉に向かい,他方は脊髄に向かって末梢組織の痛みの情報を脊髄後角(蝶形の背中半分)に伝える.この痛みを伝える神経は,一次求心性線維と呼ばれ2種類ある.後根神経節の中型ニューロンから出る有髄のAδ線維は侵害情報をすばやく伝達し,痛みの部位,強さを知覚して反射行動などを起こす警告となるのに対し,小型ニューロンから出る無髄のC線維は遅れてじわっとくる痛みを伝達する.
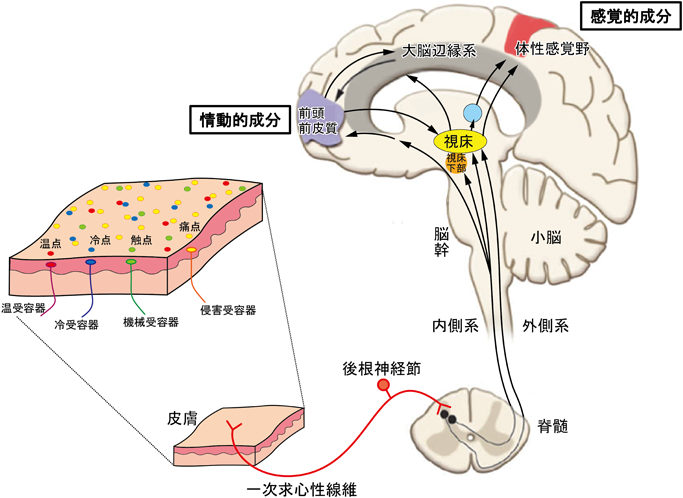
皮下の受容器で受容した感覚情報は後根神経節から出る一次求心性線維を介して脊髄を経て大脳に伝えられる.痛みには外側系を介する感覚的成分と内側系を介する情動的成分がある.
体性感覚の脳への伝達経路は脊髄後角のニューロンから出た神経線維が脊髄の正中線を越えて反対側を上行し,脳幹,視床を経て体性感覚野で感覚受容を行う外側系と痛みに伴う情動認知に関わる内側系に分けられる(図1).体性感覚,視覚,聴覚,味覚の感覚受容の入り口は視床であり,大脳皮質の感覚野の一つ,体性感覚野で情報が処理される.痛みの情報は視床下部にも送られ,発汗や動悸など自律神経系の反応を引き起こす.痛みが長引くと恐怖や情動の表現に関与する扁桃体,記憶形成に関係する海馬などの大脳辺縁系や意思決定と運動行動の制御をする前頭前皮質をはじめ複雑な脳の統合機能に関係する連合野と呼ばれる部位の関与が大きくなる.昔から「痛みをがまんする」,「痛みを分かちあう」,「痛がり」という言葉で痛みが使われているように,人によって,そのときの状況によって,痛みの感じ方はさまざまで主観的である.「各個人は,生涯の早い時期の損傷に関連した経験を通じて,この言葉をどのように使うかを学習している」と痛みの定義に注釈されるように,痛覚の情動的成分は各個人の経験・体験に依存し,記憶学習する脳の可塑的変化といえる.
痛みの基礎研究では,さまざまな疼痛モデルの系が確立し,末梢組織から脊髄,大脳と体系的に解析できる.しかも,機能分子から動物行動まで何か月もの長期間にわたって観察できる特長がある.一方,fMRIをはじめとする脳のイメージング技術の進歩により,ヒトの脳の活動部位やネットワークが可視化できるようになり,ヒトの情動的要因に影響される複雑な痛みの行動に関わる大脳の部位の理解が急速に進んでいる.「臨床的事実は基礎の実験事実よりも何百倍も正しい」とある臨床家が述べているように,痛みの研究は患者の痛みの治療に反映するという明確なゴールがあり,痛みの画期的な治療薬や根治的療法が次々と開発されている.本稿では,痛みの発生・維持機構の研究を通じて明らかになった,タンパク質分子の中に埋め込まれた構造と機能の関係と生体の精緻な仕組みに焦点を当てて解説する.
M.G. Blixは温,冷,触,痛の刺激によく応答する点が皮膚に分布していることを見つけ,感覚点と名づけた1).それらが温点,冷点,圧点,痛点で,熱刺激,圧や触などの刺激を電気信号に変換するのが,皮下にある感覚受容器である(図1).身体に危害が及ばない体性感覚の情報の多くは,すぐに受容器から脊髄への信号が送られなくなる(順応する)か,無意識下に処理されている.熱いお湯につかると慣れていい湯加減と感じる場合と熱くて痛くなり耐えられなくなって浴槽から飛び出す場合があるように,皮膚を押さえる圧刺激も度を超えると痛みに変わることを誰しも経験している.痛みを引き起こす強い刺激は侵害刺激,受容器は侵害受容器と呼ばれ,熱,機械的,化学的の三つに分類される.
スウェーデンの神経学者Å. Vallboは,手のひらに30°Cから下降あるいは上昇する温度刺激を加え,正中神経の単一の一次求心性神経の電気信号を記録し(図2A),温,冷刺激に反応し順応するものと,順応しないものの少なくとも4種類の神経活動を記録した(図2B).熱の侵害刺激は42°C付近で急に神経活動が活発になり,刺激がなくなるまでその活動は持続する.触刺激は触る,触られるだけでなく,引っぱり,圧迫,振動などを含み,機械的刺激と呼ばれる.19世紀のイタリアやドイツの解剖組織学者の名前がつけられたマイスネル小体,メルケル細胞,パチニ小体,ルフィニ終末などの構造物が圧や触受容器として表皮と真皮の境界あるいは真皮に存在する.手のひらに機械的刺激を加えて感覚神経の電気信号を記録すると,冷温受容器と同じように,これら四つの触受容器は持続的な触刺激に対してすぐ反応しなくなる性質(速順応性,RA)とゆっくり順応する性質(遅順応性,SA)を持つ電気信号への変換器であることがわかる(図2C).速順応性受容器は物の動きや振動を検知し,遅順応性受容器は物体からの圧力や形を検知する.毛の根元にも機械受容器があり,毛の動きで風を感じることができる.指を切ったときや机の角にぶつけたときに痛みを感じる機械的侵害受容器は熱の侵害受容器と同じように表皮の自由神経終末にあり,順応しない.
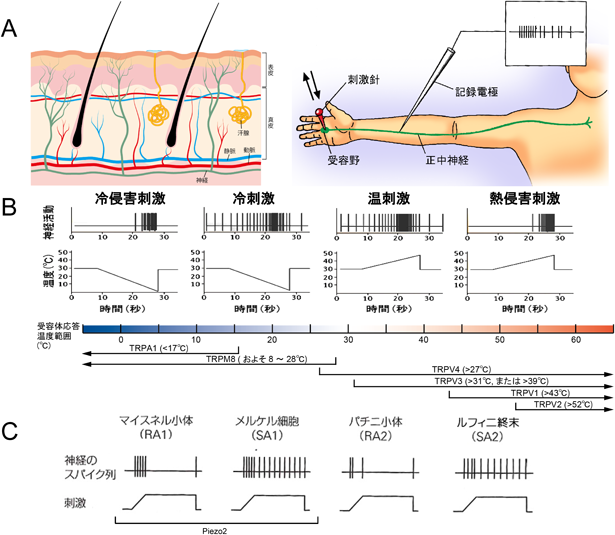
(A)皮膚の構造と単一の一次求心性線維の活動電位の記録方法.(B, C)温度刺激(B)と触刺激(C)に応答する四つの神経活動パターンと温度刺激に反応する受容器タンパク質.RA:速順応型,SA:遅順応型.
動物の新生仔にカプサイシン処理2)を行うと後根神経節の小型ニューロンが消失し,無痛状態になることから,カプサイシンが発痛物質として注目を集めるようになった.1997年にカリフォルニア大学サンフランシスコ校のD. Julius3)の研究グループは後根神経節から調製したcDNAライブラリーを培養細胞に発現させて,カプサイシンの細胞内Ca2+濃度の上昇作用を指標として,カプサイシン受容体遺伝子のクローニングに成功した.カプサイシン受容体は6回膜を貫通する構造を持ち,4分子でイオンチャネル活性を示す(図3A).カプサイシン受容体はその遺伝子の塩基配列の相同性からショウジョウバエの眼の光受容器異常変異体の原因遺伝子として同定されたtrp遺伝子のスーパーファミリーの一員であることがわかった.バニリル基を含む化学構造を持つカプサイシンにより活性化されるサブファミリーで最初に見つけられたタンパク質であることから,現在はTRPV1と呼ばれている.TRPV1は後根神経節や三叉神経節に強く発現し,カプサイシンに対するTRPV1の反応は,後根神経節ニューロンでみられたのとよく似た反応を示した.
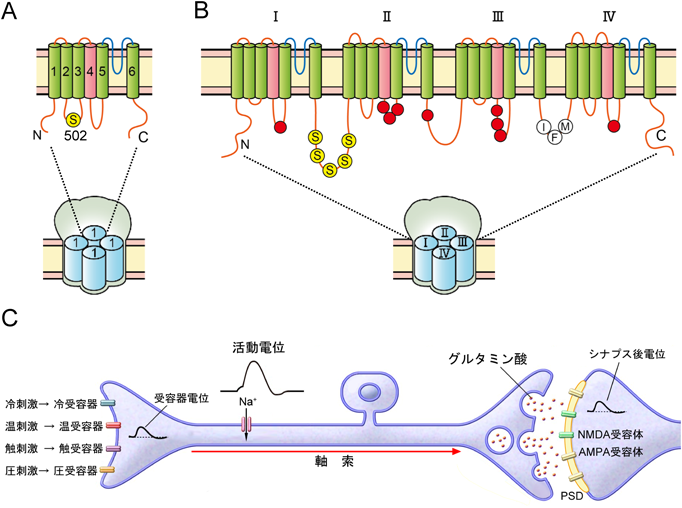
(A, B)熱侵害受容器TRPV1(A)と電位依存性Na+チャネルNav1(B)の二次構造.TRPV1は6セグメントからなる.Nav1はTRPV1と同じ二次構造のドメインI~IVでチャネルを形成する.第4セグメント(桃色)は電位センサー,第5と第6セグメントをつなぐループ(青色)はイオン選択性フィルター,第3と第4ドメインをつなぐ細胞内ループのアミノ酸配列IFMはチャネルの不活性化に関与する.Sはリン酸化部位,赤丸は遺伝性有痛症患者家系の点変異の部位.(C)一次求心性線維の神経活動に伴う電位変化.PSD:シナプス後膜肥厚.
TRPV1はカプサイシンに反応するだけでなく,43°C以上の温度で活性化されたことから熱の侵害受容器であることがわかり,痛みの生化学的研究が始まった.さらに,生体のpHは中性の7.4付近にあるが,炎症や虚血時には酸性側に傾き,TRPV1が活性化される4).TRPV1は消化管にも発現していることから,虚血時に産生される酸性物質や胃酸などの化学的な侵害受容器として内臓痛への関与も示唆されているが,機械的侵害受容器ではない.
Trpv1−/−マウスの後根神経節ニューロンの中に55°C付近で反応する細胞がごく少数みられた.Juliusの研究グループはTRPV1より高温で反応する熱侵害受容器があることに気づき,遺伝情報をコンピュータでサーチし,ラット脳からTRPV1と類似のタンパク質TRPV2を見つけた5).その後,TRPVサブファミリーはTRPV6までクローニングされている.TRPV1が主に無髄のC線維を出す後根神経節の小型ニューロンに発現しているのに対し,TRPV2は有髄のAδ線維を出す中型のニューロンに発現している.TRPV1は50°C以下の侵害刺激や炎症時の熱に対する痛覚過敏反応,TRPV2は熱いものに触った手の反射に関与し,TRPV3とTRPV4は30°C付近の温度の感知を担うと考えられる(図2B).
Juliusの研究グループは温受容器だけでなく,冷受容器もtrp遺伝子の仲間だと考えた.後根神経節のcDNAライブラリーからカプサイシンに応答するTrpv1をクローニングしたのと同じ手法を用いて,2002年に三叉神経節のcDNAライブラリーから爽快感を呼び起こすメントールに応答する遺伝子Trpm8のクローニングに成功した6).TRPM8も非選択性陽イオンチャネルである.実際,TRPM8は小型のニューロンに発現し,温度を下降させると23°C付近で活性化される冷受容器であった.2003年にはノバルティスの研究者グループにより,同じ手法で温度を下降させると17°C付近で活性化されるTrpa1がクローニング7)され,図2Bの温度刺激に対する神経活動に対応する温冷受容器がすべて同定されている.
興味深いことに,TRPV1がトウガラシの主成分カプサイシン,TRPM8はハッカの主成分メントール,TRPA1は生姜,わさびや大根で活性化されることから,温冷受容器がヒトの嗜好を形作っている.痛覚と同じように,味覚はおいしい味や店を記憶する一方,腐ったものは瞬時に吐き出すように生命の維持に大切である.温冷感覚と味覚が共通の感覚受容器を介するだけでなく,音,光が片頭痛患者の痛みの強度に影響をするのは,視床が感覚情報の大脳への入り口になっているからといえる.
触刺激の受容器は低閾値機械受容器,機械的な侵害刺激の受容器は高閾値機械受容器と呼ばれる.1997年に熱侵害受容器TRPV1が同定されたあと,15年以上経過してから2014年にメルケル細胞の触受容器としてPiezo2が同定された.この間,さまざまな分子が機械受容器の候補にあげられては消えた.低閾値機械受容器の発見は非常に単純なアイデアと力技からであった.2011年サンディエゴ郊外のラホヤにあるスクリプス研究所のグループが神経芽腫の株細胞に機械的刺激を加えて活性化されるチャネルの候補分子を二つ,DNAマイクロアレイで探し出すことに成功した.圧刺激で活性化されるこれらのチャネルはギリシャ語の圧“πιεση”(píesi)という意味からPiezo1, Piezo2と名づけられた8).
Piezo2は後根神経節に多く発現することから,2014年コロンビア大学の研究グループは皮膚のタッチドームのメルケル細胞9),シンシナチ大学の研究グループはラットの口ひげにあるメルケル細胞10)を用いてPiezo2が遅順応の触受容器である可能性を示した.
Piezo2は有毛部,無毛部どちらにも発現し,メルケル細胞だけでなくマイスネル小体,細いタイプの毛包の槍状終末,毛根を取り巻く環状の神経線維,後根神経節に広く存在することが明らかとなった.Piezo2 −/−マウスは周産期に死亡するので,コンディショナル遺伝子欠損マウスが作製され,Piezo2の機能が調べられた11).野生型マウスとPiezo2遺伝子欠損マウスで動物の行動を調べると,遺伝子欠損マウスは,綿棒で軽く触っても逃げず,弱い触刺激を加えても反応しないので,Piezo2は触受容器といえるが,高閾値機械受容器ではないようである.
生命における触刺激の重要性からPiezoチャネルは植物,粘菌,アメーバ,線虫から動物に至るまで相同遺伝子が存在するが,Piezo1とPiezo2の二つしかなく,trpチャネルのような遺伝子ファミリーを形成していない.したがって,熱刺激の神経活動に対応する少なくとも4種類の温冷受容器タンパク質が同定されたが,現在のところ,触刺激の神経活動(図2C)に対応する低閾値機械受容器タンパク質はPiezo2のみである.Piezo2タンパク質は24~39回細胞膜を出たり入ったりする巨大分子であるが,Piezoの二次構造からイオンチャネルが作られるのか,作られるとしたらどのような構造をしているのかよくわかっていない.また,機械エネルギーをどのように電気エネルギーに変換するのかもわかっていない.
Piezo2タンパク質は,四肢や頭部の位置,姿勢,動きに関する固有知覚に重要な筋紡錘やゴルジ腱器官にも発現している.マウスの尻尾を持ってぶら下げると,正常なマウスはパラシュートで落下するときのように指をのばし,四肢を広げる姿勢になる.筋紡錘に発現するPiezo2を欠損させたコンディショナル遺伝子欠損マウスでは,四肢を縮めたり,交叉したり,指をまるめたり異常な動きをとる.正常の筋紡錘でみられるI群の求心性線維の活動電位は筋肉の伸張に伴って増加するが,Piezo2のコンディショナル遺伝子欠損マウスの筋肉では活動電位はないか非常に弱く,伸張させてもあまり増加しなかった12).これらのことから,触刺激と固有知覚で感知される機械的なひずみはどちらもPiezo2が関与していることがわかり,痛みを生じない低閾値の機械的受容の全体像が明らかになりつつある.最近,Gordon症候群と呼ばれる四肢の関節の弯曲を特徴とするまれな先天性異常がPiezo2の遺伝子の異常によることが報告された13).
皮下に張り巡らされている末梢神経の終末上にある感覚受容器は,熱刺激の熱エネルギーや触刺激や圧刺激による組織の引っぱりやへこみで生じる機械的エネルギーを電気エネルギーに変換して,すばやくその体性感覚を最初の中継地である脊髄に伝えている(図1).後根神経節ニューロンや自律神経節のニューロンだけでなく,脳脊髄のニューロンが運ぶ情報は多種多様であるが,どんな種類のニューロンも軸索の細胞膜をはさむ電位変化により生じる活動電位により神経伝達が行われるところは共通である.
一次求心性線維の皮下の神経終末では冷,温,触,圧刺激により皮下の感覚受容器が活性化され,刺激によってある程度以上の大きさの脱分極が生じると神経終末に受容器電位が生じる.閾値に達すると,神経終末につながる一次求心性線維上のNa+チャネルがまず活性化され,脱分極して活動電位が生じる.一次求心性線維の神経終末近くで生じた脱分極は,ドミノ倒しのように,隣接部分の脱分極と活動電位の発生が繰り返され,皮膚での感覚情報が一方向性に脊髄に伝達される(図3C).
京都大学の沼正作教授の研究グループは電位依存性Na+チャネルが多く存在する電気ウナギElectrophorus electricusの発電器官に着目し,1984年に電気ウナギのNa+チャネル遺伝子のクローニングに成功した14).このタンパク質は1820個のアミノ酸からなる途方もない大きな分子で,さらに,1986年にラット脳に存在する3種類の電位依存性Na+チャネルの二つ,Nav1.1とNav1.2のαサブユニットの完全な塩基配列の決定15),1988年にNav1.3のαサブユニットの塩基配列の決定に成功した.ラット脳のNa+チャネルは約2000個のアミノ酸からなる巨大タンパク質で,Nav1.2のcDNAから合成したmRNAをアフリカツメガエルに発現させると,記録される電流は興奮性細胞のNa+チャネルと類似したチャネル活性を示すことが確認された16).
Nav1のαサブユニットは一つの長いポリペプチドから形成されており,分子量26万の非常に大きな膜タンパク質で,構造がよく似たドメインが四つあり,1分子でチャネルを形成する(図3B).そして,A. HodgkinとA. Huxlayが電位固定装置を用いた実験から提唱したNa+チャネルの三つの特性—Na+イオンの選択性,電位依存的な活性化,チャネルの急速な不活性化—がすべてNav1の一次構造のアミノ酸配列の中に組み込まれていることがmRNAの機能的発現で解明された.沼研究室での主要なイオンチャネルのクローニングはイオンチャネルの研究に革命をもたらすとともに,タンパク質科学から遺伝子工学への時代の移行を印象づけた.興味深いことに,1997年に同定されたTRPV1はNav1のドメインと同じ二次構造(図3A, B)を持ち,4分子で1チャネルを形成する.
Nav1はNav1.1~Nav1.9の9種類あり,後根神経節には,Nav1.1, Nav1.6, Nav1.7, Nav1.8とNav1.9の5種類のNav1が発現している.Nav1.7, Nav1.8とNav1.9が主に小型ニューロンに発現するのに対し,Nav1.6は後根神経節のすべてのニューロンに,しかも強く発現している.Nav1.7とNav1.8は神経突起の先端,すなわち自由神経終末に多く発現し,Nav1.8は痛覚過敏に関係することが知られている.
パキスタン北部の三つの家族で生まれつき痛みを感じない4歳から14歳の男児4人,女児2人が見つかった.彼らは生まれてこのかた,体のどの部位にも痛みを感じたことがなかったが,触や圧覚,温度覚や固有知覚は保たれており,知能も正常であった.ユニバーシティ・カレッジ・ロンドンのJ. N. Woodをはじめとする英国の研究者グループは患者のDNAを解析してNav1.7をコードする遺伝子SCN9Aの異常が原因であることを見つけた17).Nav1.7は1977個のアミノ酸からなる大きな分子である.パキスタンの先天性無痛症患者の3家系では,SCN9A遺伝子の点変異でストップコドンに変わり,正常なNav1.7タンパク質が作られず,無痛症になっていた.その後の調査で,7か国の9家系でNav1.7の異常により先天性無痛症になる患者がいることが明らかとなった.
2004年に中国人の家系の中に,運動や少し手足が温められることが引き金となって,両方の手足の焼けつくような痛みと皮膚が赤く熱を帯びる発作が数分から数時間続き,冷やすことで軽くなる遺伝性肢端紅痛症の患者が報告18)された.この家系はNav1.7の858番目のロイシンがヒスチジンに変わる点変異の優性遺伝である.この家系以外にもNav1.7の点変異による遺伝性有痛症の患者が見つかっている(図3Bの赤丸).原因が不明の有痛性ニューロパシーの30%はNav1.7のイオンチャネルの活性化によると報告されており,ヒトの疾患から,Nav1の中でNav1.7が痛みの発生,神経伝達に重要であることがわかる.
冷たいものを飲んで歯がしみる,海水浴の後温い風呂が熱い,捻挫をして足が腫れて歩くのも触るのも痛いというように,皮下の侵害受容器から脊髄への入力が増えることにより,痛みが増強する.このような歯や皮下で起こっている変化を末梢性感作と呼ぶ.侵害受容器には,順応性がないため,刺激に敏感になり,受容器電位が持続して発生すると痛みが持続することになる.受容器電位の発生がなくても,上に述べた遺伝性有痛症患者のように,Nav1.7の活動電位が発生しやすくなる場合も痛みが生じる.末梢性感作は,次の実験から侵害受容器あるいはNav1のリン酸化で説明できる.
TRPV1がリン酸化されていない場合には,温度の閾値が42°Cで初めてチャネルが開き,受容器電位が発生する.TRPV1の第2と第3セグメントの間の502番目とC末端付近にある800番目のセリン残基(図3A)がプロテインキナーゼCでリン酸化されると,TRPV1のチャネルの性質が変化して30°C付近からチャネルが開くようになり,低い温度から侵害情報として脊髄に伝達される19).Nav1のドメインIとIIの間の細胞内ループにセリン残基が五つ存在し(図3B),プロテインキナーゼAでリン酸化されると,大きな内向き電流がみられるようになった20).このことから,Nav1はリン酸化により自発的に活動電位を発生するようになると考えられる.
このように,我々の体の機能は,電灯のスイッチのように,意外なほど簡単な仕組みでチャネル活性がオン・オフされている.遺伝性有痛症の患者の症状が一つのアミノ酸の点変異で生じるように,TRPV1とNav1のリン酸化はこれらの機能を変化させるのに十分であることを示しており,タンパク質のリン酸化は生後に起こる可逆的なアミノ酸の点変異といえる.
プロスタノイドは,炎症に伴って細胞膜から切り出されたアラキドン酸からプロスタグランジン合成酵素(COX)により5種類が合成される.そのプロスタノイドの中で,炎症時の発熱や痛みに中心的な役割をするのがプロスタグランジンE2(PGE2)である.プロスタノイド受容体はいずれもGタンパク質共役型受容体であるが,PGE2だけがEP1~EP4の4種類のEP受容体サブタイプがあり,それぞれのサブタイプを介してプロテインキナーゼAやプロテインキナーゼCなどのキナーゼを活性化して,末梢組織での痛覚過敏反応に関与する.炎症を抑えるステロイドホルモンはアラキドン酸の遊離を抑制し,アスピリンはCOXを阻害して鎮痛効果を発揮する.1899年に市場に登場したアスピリンは,100年以上にわたり鎮痛薬のベストセラーで,アスピリンなどの鎮痛薬は,ステロイドと区別するために非ステロイド性消炎鎮痛薬と呼ばれる.
このように,PGE2が炎症部位で末梢性感作に作用すると考えられていたが,1992年に我々は脊髄腔内に投与したPGF2α,次いでPGE2が触刺激による痛み(アロディニア)を誘発すること21, 22)を発見し,これから述べる脊髄における痛みの発現・維持機構の研究がスタートした.その当時,帯状疱疹後神経痛のような神経障害性疼痛では,帯状疱疹が治癒した後,アロディニアが生じることは臨床症状として知られていたが,アロディニアという言葉はほとんど使われていなかった.そして,長い間,神経障害性疼痛は脊髄の神経回路の再構成23)や脊髄ニューロンの細胞死による器質的変化で生じるため難治性と考えられた.
末梢からの侵害性,非侵害性入力に対して脊髄ニューロンの発火頻度が上昇して脳に過剰な感覚情報が伝えられる変化は中枢性感作と呼ばれ,慢性痛の重要な基盤となる.痛みの感受性を増す中枢性感作の基礎にはシナプス伝達の促通があげられる.末梢性感作に伴うシナプス前終末からグルタミン酸の放出量が増加する機構に加えて,シナプス後細胞の伝達効率が増す機構がある.ニューロンの細胞体だけでなく,シナプスを形成する受け手側の樹状突起から出るスパインと呼ばれる小さな突起でもシナプス伝達の促通が生じる.
神経伝達物質としてグルタミン酸を利用している中枢神経系の興奮性シナプスは,シナプス前終末から遊離されるグルタミン酸をAMPA受容体とNMDA受容体が集合したシナプス後膜肥厚(PSD)で効率よく受け取っている(図3C).一次求心性線維を介して侵害刺激が入力した場合にAMPA受容体が活性化され,一過性の痛みを誘発するが,痛みはすぐに消える.痛みを伝える無髄の神経線維の入力を10秒~20秒の間隔で続けると,脊髄後角のシナプスでの脱分極反応が累積して,活動依存的なNMDA受容体のMg2+ブロックがはずれ,Ca2+イオンを通すNMDA受容体が活性化される.NMDA受容体のグルタミン酸に対する感受性が増加すると連続する入力の一つ一つの刺激に対して活動電位の数が増加していくとともに,電位依存性のCa2+チャネルが参加するようになり,持続する脱分極の確立に貢献する.脊髄後角ニューロンの反応が低頻度による刺激で増加する現象をワインドアップ24)と呼ぶ.刺激の強さが変わらなくても連続する熱や機械的な刺激で痛みに変わるワインドアップ現象は,持続的な熱侵害刺激の間,停止することがなく受容器電位を出し続けるTRPV1あるいはTRPV2の反応(図2B)とよく合っている.しかし,慢性痛では多くの侵害刺激が高頻度で同期して入力することはありえないので,ワインドアップによって中枢性感作が形成されるとは考えにくい.
NMDA受容体はGluN1の二つとGluN2の4種類のサブユニットGluN2A~2Dのうちの二つでイオンチャネルを形成する(図4A).GluN1はNMDA受容体のチャネル活性に必須であるので,大脳皮質,海馬,小脳をはじめ脳全体に分布している.NMDA受容体の機能的多様性はMg2+ブロックの有無やリン酸化による機能の修飾が異なるGluN2サブユニットの構成により決まる.神経障害性疼痛モデルの痛覚過敏反応や触刺激による痛み(アロディニア)の症状がNMDA受容体,中でもGluN2Bの拮抗薬で緩和されたことから,我々はNMDA受容体GluN2Bサブユニットが中枢性感作に関与することを示した25).
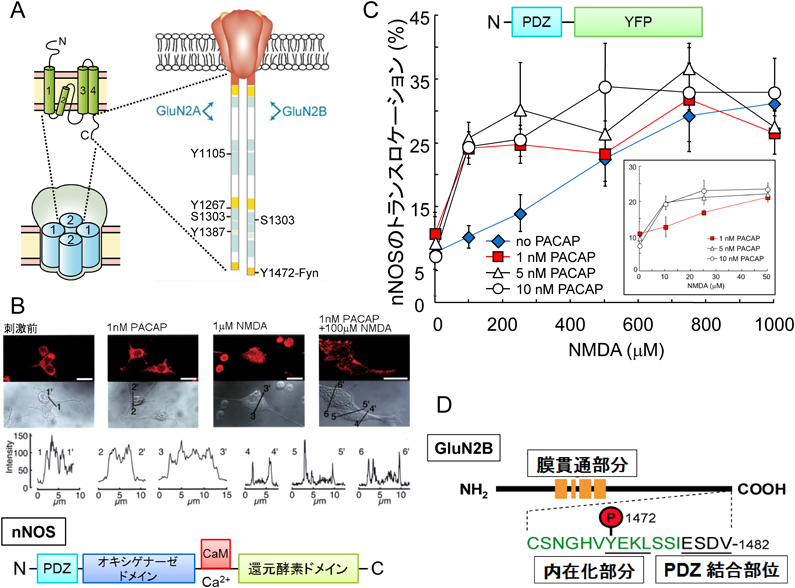
(A) NMDA受容体の二次構造とリン酸化部位.(B) nNOSの細胞内移動とタンパク質構造.(C) PACAPによるnNOSの細胞内移動に必要なNMDA濃度の低下.(D) GluN2BのY1472近傍のアミノ酸配列と機能.
GluN2AとGluN2BサブユニットはC末端側に非常に長い細胞内部分を有しており,GluN2BのC末端近傍の1472番目のチロシン残基(Y1472)はチロシンキナーゼ活性を持つSrcファミリーの一つFynの主なリン酸化部位26)で,GluN2Aにはない(図4A).GluN2Bは脊髄後角の浅い層に局在する.神経障害性疼痛モデルではY1472のリン酸化が顕著に増加し,Fyn −/−マウスでは神経障害性疼痛が生じなかった25).さらに,GluN2BのY1472を水酸基がないフェニールアラニンに置換してリン酸化できなくしたマウスでも神経障害性疼痛が生じなかったこと27)から,GluN2BサブユニットのY1472のリン酸化が神経障害性疼痛の発生に関与することが確かめられた.神経障害性疼痛モデルが完成した1週間後でもGluN2Bの拮抗薬で一過的に神経障害性疼痛が緩和されたこと25)は,神経障害性疼痛がGluN2Bのリン酸化で維持されていることを意味しており,意外であった.なぜなら,不可逆的な中枢性感作機構のため難治性と考えられていた神経障害性疼痛が,GluN2Bの脱リン酸化で和らぐ可能性を示唆したからである.
PSDの下には細胞質やスパインの突起に広がる細胞骨格タンパク質群がある.GluN2AやGluN2BのC末端側にある細胞内部分は細胞内に打ち込まれた杭のように,細胞質や細胞膜にある広範なタンパク質群との相互作用により,それぞれの適切な場所への局在が確保できる.PSDのNMDA受容体複合体28)は細胞骨格タンパク質を含む100近くのタンパク質で複雑な網目構造を形成する.そして,シナプスの活動状態によりダイナミックに解離会合を行っている.
神経損傷時には後根神経節で下垂体アデニル酸シクラーゼ活性化ポリペプチド(PACAP)の発現が誘導された29).PACAPはGタンパク質共役型受容体PAC1を介して神経伝達物質,神経調節因子,神経栄養因子のような多彩な神経作用を発揮する.PACAP−/−マウスでは脊髄後角表層で神経型一酸化窒素合成酵素(nNOS)の活性化がみられず,神経障害性疼痛が生じなかった.興味深いことに,神経株細胞で酵素タンパク質の局在を調べると,通常nNOSは細胞質に局在するが,NMDA受容体作動薬NMDAとPACAPで同時に刺激すると,nNOSは細胞質から細胞膜へ移動し(図4B),一酸化窒素を産生するようになった.このことは,NMDA受容体が活性化されるとnNOSがNMDA受容体複合体に加わることを意味する.
3種類のNOSの中で,細胞膜のNMDA受容体や足場タンパク質との会合に重要なPDZ結合ドメインがあるのはnNOSだけである(図4B).細胞内のnNOSの動きを追跡するために,PDZ結合ドメインに緑色蛍光タンパク質の変異体である黄色蛍光タンパク質をつないだ融合タンパク質(図4C)を作製した30).NMDAだけで刺激すると融合タンパク質の細胞膜への移動には細胞死を起こすほど高濃度のグルタミン酸(500 μMから1 mM)が必要であった.驚いたことに,1~10 nMの低濃度のPACAPと同時に刺激するとnNOSの移動に必要なNMDA濃度を10~50 μMまで下げることができた(図4C).シナプス終末から遊離されるグルタミン酸でnNOSが細胞膜に移動することができる濃度である.nNOSがNMDA受容体の近くに移動するのは,NMDA受容体チャネルを通って入るCa2+濃度の高い部分に移動して,効率よく一酸化窒素を合成するためと考えられる.
GluN2BサブユニットのY1472のリン酸化部位近傍には,GluN2Bを含むNMDA受容体のインターナリゼーション(内在化)に関係するアミノ酸配列YEKLと,nNOSが結合するのに特徴的なPDZ結合モチーフESDVが存在する(図4D).NMDA受容体は刺激がなくなるとスパイン内に取り込まれ,PSDから除去されるが,Y1472がリン酸化されるとGluN2BのC末端部分と足場タンパク質PSD-95が結合して,内在化が阻止される31).さらにnNOSと複合体を形成し,NMDA受容体を通って流入するCa2+がnNOSの活性化を起こす.すなわちGluN2BのY1472のリン酸化部位近傍はシナプス終末からの情報の「受容の持続」と脊髄ニューロンへの「シグナル伝達」という二つの機能が埋め込まれていることがわかった25).
脊髄腔内に投与したPGE2がアロディニアを誘発することを発見したこと22)から,我々は脊髄でのPGE2の中枢性感作に関与する作用とそのメカニズムを詳細に調べた.
神経損傷ではその異常を察知してミクロクリアが活性化され,損傷部位に集積する.Gタンパク質のGiとGsはアデニル酸シクラーゼの抑制と活性化に拮抗して正反対に働く.損傷した部位から放出されるATPはGiと共役するATP受容体P2Yを介してミクログリアを損傷部位に遊走させる(図5).我々は培養したミクログリアでEP受容体サブタイプの発現を調べ,Giと共役するEP3だけ発現しておらず,PGE2はGsと共役するEP2を介してプロテインキナーゼAを活性化して,遊走するミクログリアの動きを抑制することを明らかにした32).一酸化窒素も細胞膜をすり抜けて,ミクログリアの動きを抑制し,結果としてミクログリアは侵害情報を伝える一次求心性線維の中枢端に集まることになる.一次求心性線維の脊髄の中枢端やミクログリアで産生されるPGE2はシナプス終末のグルタミン酸(Glu)の遊離を促進した33)が,おそらくCa2+動員作用を有するEP134)を介すると考えられた.EP3は細胞骨格タンパク質アクチンに作用してnNOSの移動,活性化に関係し35),EP4はアロディニアを誘発するオピオイドペプチドN/OFQの遊離を促進させた36).このように,PGE2が4種類のEP受容体EP1~EP4を介する多彩な作用機構により脊髄の中枢性感作の維持に関与することを明らかにした.さらに,EP2がシナプス伝達に抑制的なグリシン受容体を脱抑制することも報告されている37).
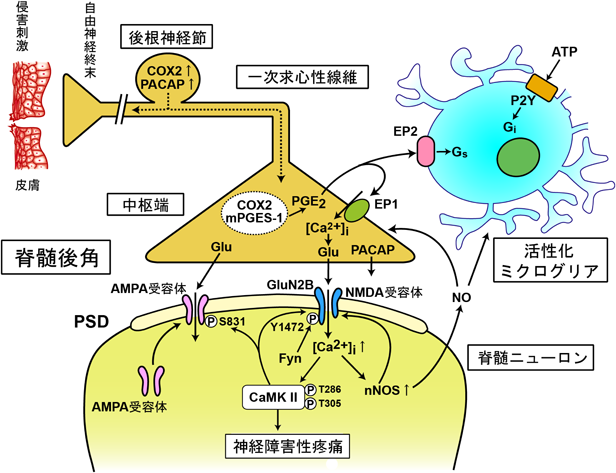
末梢神経が損傷されると,後根神経節のシクロオキシゲナーゼ2 (COX2),下垂体アデニル酸シクラーゼ活性化ポリペプチド(PACAP)をはじめとするさまざまな遺伝子の発現に変化が生じ,神経終末に運ばれる.遊離されるグルタミン酸(Glu)の増加により,AMPA受容体だけでなくNMDA受容体も活性化され,流入するCa2+によりnNOSやCa2+/カルモジュリンキナーゼII(CaMKII)など脊髄ニューロンの細胞内シグナルカスケードが活性化される.活性化されたミクログリアが集まってさまざまな生理活性物質が遊離され,脊髄後角は反応の場となり,中枢性感作が生じる.mPGES-1:膜型プロスタグランジンE合成酵素-1,NO:一酸化窒素.
健康なときは,侵害刺激の神経伝達は細胞膜の活動電位とシナプスでの瞬間的なグルタミン酸の放出により,静止膜電位近くでAMPA受容体が活性化され,一方向に伝達され,痛みは一過性である.慢性痛では末梢性感作や中枢性感作が生じ,痛みが持続する.図5は我々が明らかにした脊髄で生じる機能的な中枢性感作のまとめである.炎症や神経損傷で末梢組織から持続的に侵害情報が送られてくると,PSD上のAMPA受容体,NMDA受容体のリン酸化により,チャネル活性を上昇させるとともに,グルタミン酸受容体の移動を調節して受容体の数を増やし,活性化の時間を延長させる.NMDA受容体はCa2+イオンをよく通すので,シナプス後ニューロンの細胞内の遊離Ca2+濃度の上昇により,nNOSやCa2+/カルモジュリンキナーゼII(CaMKII)を含むさまざまなCa2+依存性の酵素が活性化し,シナプスの伝達効率が増加する.Na+イオンやCa2+イオンと異なり,一酸化窒素は細胞膜を自由に透過できることから,脊髄後角のニューロンから逆行して一次求心性神経のシナプス終末やミクログリアに作用することができる.壊れたニューロンからATPが漏れ出し,ミクログリアが集積するとサイトカインやPGE2などの生理活性物質が放出され,神経伝達の一方向性が崩れる.このようにして,脊髄後角は遊離された生理活性物質やミクログリアが集積して反応の場となり,炎症部位での末梢性感作のように,長時間持続する中枢性感作を引き起こす.しかし,脊髄での中枢性感作には非常に多くの分子,細胞が関与するので,図5は中枢性感作の発生機序のそのほんの一部を示したにすぎない.
これまで述べてきたように,熱の侵害受容器はTRPV1チャネル,触受容器はPiezo2チャネル,神経伝達に不可欠な分子はNav1,シナプス伝達におけるAMPA型とNMDA型グルタミン酸受容体と,痛みの発生や持続に関係する重要なタンパク質はいずれもイオンチャネルである(図3, 4A).TRPV1とNMDA受容体はいずれも非選択的陽イオンチャネルで,リン酸化によりチャネル活性が増強され,Ca2+の流入により細胞内カスケードを活性化させて末梢性感作,中枢性感作に関係している.Nav1.7の欠損は先天性無痛症になり,Nav1.7のアミノ酸の点変異は遺伝性有痛症になる.これらのことから,痛みはチャネル病といえる.
複合性局所疼痛症候群(カウザルギー)は,主に外傷後に起こる四肢の遷延する代表的な難治性の神経障害性疼痛である.これまで,カウザルギーの中枢性感作症状は,一次求心性線維の脊髄での神経回路の再構築や抑制性ニューロンの細胞死など器質的変化により難治性であると説明されてきた.そして,末梢性感作と中枢性感作の両方が密接に関係するため,1864年にカウザルギーの最初の報告がなされて以来,140年間治らないとされてきたが,2005年に生体内神経再生によりカウザルギーの根治的治療に成功したことが初めて報告された38).それ以降,数百例の患者の多くで,器質的な中枢性感作説で説明されてきたさまざまな症状,長期間持続した症状が末梢神経再生で完全に消失したことから,中枢性感作は器質的変化ではなく機能的変化によることが臨床的に実証されている.図5の機能的かつ可逆的な中枢性感作機構は,末梢性の神経障害性疼痛は原因疾患が治療できれば痛みが消失することと合致している.
神経再生には,細胞,液性因子と再生の場や環境が大きく影響する.脊髄損傷のような中枢神経系の神経再生では,iPS細胞を用いた再生医療が注目を浴びているが,阻害因子や脊髄損傷後の限られた治療時期などの要因により神経再生させること,さらに機能回復をさせることは容易でない.一方,カウザルギーなどの神経損傷後の神経障害性疼痛は,末梢神経系の旺盛な神経再生能があだとなり,神経再生の場・環境が適切でないと正しく標的臓器に到達できないために生じるといえる.末梢神経では切断後,運動ニューロンや感覚ニューロンとつながっている近位側は切断後も軸索構造が維持されるが,遠位側の神経線維は断片化,除去され,再生した神経が通る神経鞘が残ることが知られている(Wallerian変性).次に述べる我々の末梢神経再生の最近の研究によりニューロンから再生した軸索は神経鞘を通り,標的臓器に到達する神経再生のメカニズムは非常に精緻にできていることが明らかとなった.
これまでの末梢神経再生の研究では,再生のある時期に神経組織を取り出し,その組織標本から神経再生の状況を判断していたため,得られる神経再生の情報は断片的で,個体差を考慮しなければならなかった.我々が確立した神経組織に選択的に蛍光タンパク質を発現するthy1-YFPマウスを用いる坐骨神経切断−再生モデル(図6A)は,大腿部の坐骨神経を切断し,切断端を5 mmの間隙を残してシリコンチューブでつなぐシンプルなモデルである39).このモデルで,生きたマウスの神経再生過程を同一個体で経時的に,何か月もの長期間にわたって追跡すると,シリコンチューブ間の0.5 cmを再生するのに3週間かかるのに対し,再生神経が切断神経の遠位端に到達した後,足先(約4 cm)や腓腹筋に到達する速度は驚くほど速かった(図6B).この興味深い現象の答えは次に述べる意図しない実験から得られた.
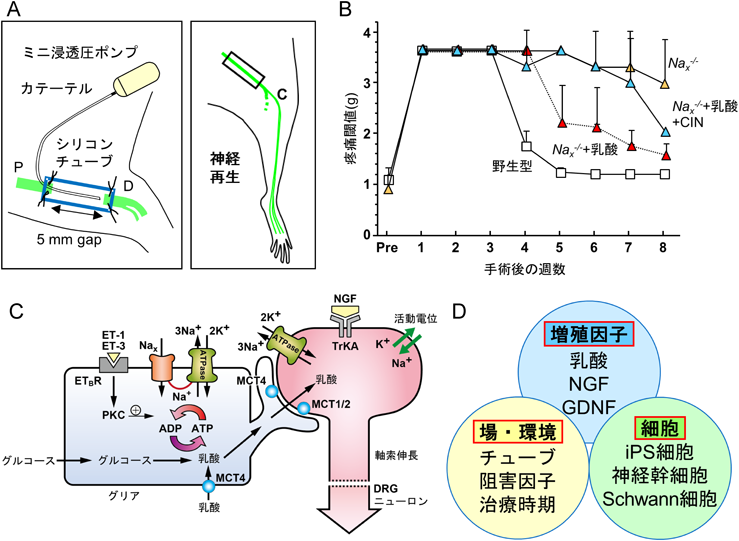
(A) thy1-YFPマウスを用いる坐骨神経切断−再生モデルの模式図.(B) Nax遺伝子欠損マウス(Nax−/−)の神経再生の遅延.(C)末梢神経の再生メカニズムの想定図.ET:エンドセリン.(D)神経再生の三つの要素.
Nav1遺伝子ファミリーに属するNaxは電位依存性Na+チャネル活性がなく,160 mM付近のNa+濃度で活性化されるNa+濃度依存性Na+チャネルであること,血中のNa+濃度を感知して摂水行動に関与することが野田昌晴博士のグループにより明らかにされた40).末梢神経系ではNaxが後根神経節や末梢神経のSchwann細胞に強く発現している.Nax−/−マウスで坐骨神経切断−再生モデルを作製したところ,予想外なことに,神経再生が大幅に遅れ,ミニポンプで4週間乳酸を持続注入すると,野生型よりわずか1週間遅れで神経再生がみられた(図6B)41).乳酸は細胞膜上のモノカルボン酸輸送体(MCT)を介して輸送されるので,その阻害剤CINを乳酸と同時に投与すると,乳酸の促進効果がなくなり,Nax−/−マウスと同じように神経再生が大幅に遅れた.
Naxチャネルが最大値の50%活性化されるNa+濃度は161 mMであるが,血管収縮因子のエンドセリン-3を1 nM添加すると活性化に必要なNa+濃度が正常なNa+濃度133 mMまで下がる40).エンドセリンは図4Cに示したPACAPがnNOSの移動に必要なNMDA濃度を下げるのと同じ役割をしている.さらに,NaxのC末端部分にNa+/K+-ATPaseとタンパク質会合するアミノ酸配列が埋め込まれていることが報告されている42).このような報告と再生神経の近くに太い血管がみられることを勘案すると,生体における末梢神経再生の精緻なメカニズムが浮かび上がってきた(図6C).再生血管より遊離したエンドセリンは正常なNa+濃度でNaxを開かせる.活性化されたNaxを通って流入したNa+イオンをくみ出すために,Naxとタンパク質間相互作用をするNa+/K+-ATPaseが活性化し,ATPを消費する.グリア(Schwann細胞)はATPを供給するため,グルコースの取り込みや解糖系を促進させ,乳酸を産生する.産生された乳酸はMCTのサブタイプMCT4を介してニューロンや軸索に運ばれ,エネルギー源として神経再生に用いられるというニューロン−グリア間の連携が浮かび上がってきた43).
モデルで切断する坐骨神経は運動神経と感覚神経の混合神経であるが,神経再生過程で運動神経と感覚神経が誤った標的組織,筋肉と皮膚,に投射する可能性はないかほとんどなかった.その理由は,皮膚からは神経成長因子(NGF),筋肉からはグリア細胞株由来神経栄養因子(GDNF)といった異なる神経栄養因子が標的臓器から分泌される.それらの受容体を発現する神経突起が神経栄養因子を取り込み,逆行性輸送されたニューロンが生存し,軸索を伸長させ標的臓器に神経線維を投射するというLevi-Montalciniの説が,発生だけでなく神経再生でも働くためである.これらのことから,3週間かけてシリコンチューブの遠位側に到達した再生神経は,神経鞘の内部を形成するSchwann細胞から供給される乳酸をエネルギー源として再生神経の伸長が加速され,標的臓器から遊離される神経栄養因子により正しく標的臓器に投射されると考えられる.末梢神経系では神経栄養因子だけでなく,新たにSchwann細胞と乳酸が神経障害性疼痛の生体再生医療の標的に加わった(図6D).
マウスは口と両耳の穴の3点で頭を固定できるので,生きたマウスの脳のニューロンや神経線維の形態変化を顕微鏡下に観察することは必ずしも難しくない.一方,脳脊髄液に浮かぶ脊髄は呼吸による横隔膜の上下運動や心臓の拍動の影響を受けるので非常に困難であったが,最近,我々は生きたマウスの脊髄での形態変化を長時間観察することに成功した44).神経組織に選択的に黄色蛍光タンパク質を発現するthy1-YFPマウスの後肢に起炎物質を注入して,多光子顕微鏡を用いて脊髄後角ニューロンとその神経突起の変化を観察すると,注入後30分ですでにスパインの数が増え,神経線維のふくらみもみられた.シナプスの数を計算すると,炎症に伴って30分後からシナプス数が1.3~1.4倍に増加し,その増加は6時間の実験期間中観察された.AMPA受容体やNMDA受容体の拮抗薬,電位依存性Ca2+チャネル阻害薬により,シナプス数の増加がみられなくなった.これらのイメージングの結果から,炎症に伴って脊髄ニューロンのシナプスでの伝達効率の上昇,細胞内シグナルカスケードの活性化(図5)に引き続いて,形態変化も起こっていることがわかった.記憶学習に関与する海馬の長期増強でシナプスでの伝達効率が増えるだけでなくスパインの数が増えることがよく知られており,脊髄での中枢性感作と記憶学習に関連する海馬の長期増強には,メカニズムおよび関与する分子の共通点が少なくない24).GluN2Bのリン酸化は海馬の長期増強のときに増大すること26),そして,その重要性は神経障害性疼痛の持続だけでなく,アルツハイマー病の記憶障害にも関係することが報告されている.
脳幹の中脳水道周囲灰白質(PAG)(図7A)を刺激すると,脊髄からの侵害情報の脳への出力が抑制されること,少量のモルヒネをPAGに注入すると鎮痛作用が生じることから,侵害情報の伝達を強力に修飾するPAGから脊髄への経路は下行性疼痛抑制系と名づけられた.阿片から名づけられたオピオイド受容体は,モルヒネの鎮痛作用と一致して,視床,PAGや脊髄に分布している.興味深いことに,内在性オピオイドペプチドのβ-エンドルフィンはストレスに応答する副腎皮質刺激ホルモンと同じ前駆体プロオピオメラノコルチンに由来し45),痛みとストレスが密接に関係することが理解できる.
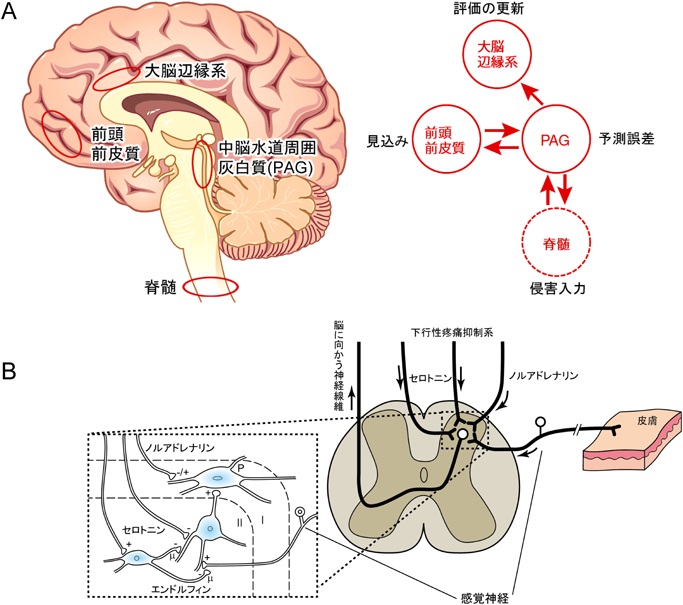
(A)侵害情報は内側系を介して大脳辺縁系や前頭前皮質に送られ情動反応を引き起こすだけでなく,PAGにフィードバックされる情報は大脳辺縁系と脊髄に送られる.(B)脳幹部から出るノルアドレナリンとセロトニン作動性線維が下行性疼痛抑制系で,内在性オピオイドのエンドルフィンと協調して脳に侵害情報を送る脊髄ニューロンの閾値を調節する.
脊髄や大脳皮質はニューロンが集まって層構造で神経回路網を作り上げているのに対して,脳幹の中心部の神経回路網はニューロンからなる神経核を形作らず,網目状になり神経線維束の白質と混在していることから,脳幹網様体賦活系と呼ばれる.脳幹は入力される感覚情報を受け取り意識や覚醒レベルの調節を行い,多種多様な情報源からの情報を統合して,感覚,運動,自律機能へ強い影響を及ぼすように構成されている.脳幹部は,ドーパミン,セロトニン,ノルアドレナリン,ヒスタミンなどの神経伝達物質を含む細胞群とそれらから出る神経線維からなる.ノルアドレナリンを合成するニューロンが脳幹の橋の部分にあり,合成されたノルアドレナリンが大脳のすべての領域に運ばれる一方,脊髄にも運ばれている.セロトニンの神経線維はノルアドレナリンと同じように脊髄にも神経を出して下行性疼痛抑制系に関わっている(図7B).
神経伝達物質のドーパミン,ノルアドレナリンはアミノ酸の一種チロシン,セロトニンはトリプトファン,ヒスタミンはヒスチジンから簡単な酵素反応で合成される.興奮性伝達物質のグルタミン酸や抑制性伝達物質のグリシンはアミノ酸そのものである.生体に豊富に存在するアミノ酸とその代謝産物でヒトの神経活動,精神状態や意識が制御されていることを不思議に感じるとともに,気分や感情は揺れ動くものだと納得させられる.
意識は知覚,思考や行動など精神活動の多くの側面に必要だが,神経系はさまざまな入力信号からある一部の情報を抽出して他の多くを無視する.言い換えると,我々の脳は入力されるさまざまな入力情報を同時に処理できるほどの能力がない.そのため,痛みの意識的な情動認知は感覚受容とは質的に異なり,得られる入力情報の一部を抽出し処理するので,一人一人の痛みの受け止め方は主観的にならざるをえない.
痛みはどのような行動が自分に利益をもたらすのか,避けるべきなのかを判断する信号となる.I. P. Pavlovは著書『大脳半球の働きについて—条件反射学—』で「餌をもらう前に,いつも電気ショック,熱刺激や強打などの罰を与えられた犬は痛みを伴う強い侵害刺激が餌の合図であるかのように尻尾を振る」と記載している46).このような疾病利得による痛みは学習痛として知られている.これは極端な例であるが,餌をもらうといった利得の予測は脳幹部から出るドーパミンニューロンと側坐核がよく知られている.予期しないときに報酬が得られた場合にドーパミンの分泌が増え,予期したときに報酬が得られた場合にはその分泌に変化がなく,予期したときに報酬が得られない場合にはその分泌が減る応答をする.期待値と実際の報酬の差が報酬予測誤差であり,我々の日常生活でも予測誤差が生じた場合には,常に修正しながら行動している.罰に伴う痛みとそれを避ける学習は行動の意思決定に重要な影響を及ぼし,適応できない体験やストレスが慢性痛の引き金となり,避けられない多くの嫌な出来事が慢性痛を維持させる重要な要因となる.侵害刺激で侵害受容器が活性化され,侵害情報は脊髄から外側系を経て体性感覚野に伝えられて痛みを知覚する一方,内側系を経て大脳辺縁系や前頭前皮質に送られ情動反応を引き起こす(図1).しかしながら,電気ショックや殴打などの罰の予測誤差に関与する部位はよく知られていなかった.最近,熱侵害刺激を与えた場合にその罰の予測の中心がPAGにあることがヒトの脳のfMRIで明らかにされた47).そして,PAGは前頭前皮質からの痛みの予測値と脊髄から来る侵害情報の値を統合して得られた予測誤差を大脳辺縁系の前帯状回に送り,その評価が更新されるという構図が提案された(図7A).このように,PAGは大脳辺縁系や前頭前皮質に侵害情報を送り,処理された情報に基づいてこれまでの痛みの体験に上書きして記憶に反映させる一方,下行性疼痛抑制系を介して脊髄の侵害情報を脳に伝える閾値を調節するのにいい位置を占めており,PAGは痛みの制御・統合の中心にあるといえる.
これまでの研究手法では,刺激を与えて反応をみる,そして,ある仕事をしている神経活動部位をfMRIの機能的イメージングで調べるということに焦点が当てられてきた.痛みを起こさせたときに特異的なシグナルをいかに取り出すか,すなわち,安静時のランダムな自発的な脳の活動をノイズと考え,減らして特異的なシグナルの比率,S/N比をあげることに精力が注がれてきた.最近,これまでノイズと考えられてきた自発的な脳の活動が,デフォルトモードネットワークとしてとらえられて,着目されている48).
大規模な脳の固有の神経回路網を研究するなかで,安静時に脳の自発的な変動が同期する空間的パターンが浮かび上がってきた.脳が活動しているときに大脳辺縁系の後部(黄色の矢印)と一致して前頭前皮質の内側(橙色の矢印)も青くなり活動が低下している(図8A).デフォルトモードネットワークの活動の低下は,視覚,聴覚,体性感覚などの感覚情報や運動で脳の他の部位の活動が上昇するとき,すなわち作業モードのときにみられる.反対に,脳が休んでいるときにこれら二つの部位が連動して赤くなり,活動していることが示されている.fMRIの機能イメージングを折れ線グラフにして時間変化で表すと二つの部位が同期していることがよくわかる.このネットワークの機能は決して途切れることなく,活性を上げたり下げたりしている.図1に示すように,前頭前皮質は感覚情報を受け入れ,大脳辺縁系,視床下部やPAGに情報を流す部位であり,個人の人格に関わる社会的行動,気分の制御,動機づけなどに関わっている.もう一つのデフォルトモードネットワークの部位は後部帯状回である.側頭葉の外側とともに記憶の固定に関係している.このネットワークは夜に強く活動しており,昼間活動した体験をとりまとめて,毎晩リセットしていることになる.重要なことは,デフォルトモードネットワークは特定の仕事の作業と関係なく,活性が高い基底状態にあり,我々は脳の部位の活性をわずかに変化させて,作業モードの活性化に対応している.そして,驚くべきことに,大脳のエネルギー消費の多くは,安静時のデフォルトモードネットワークの自発的活動,つまりメインテナンスに使われている.
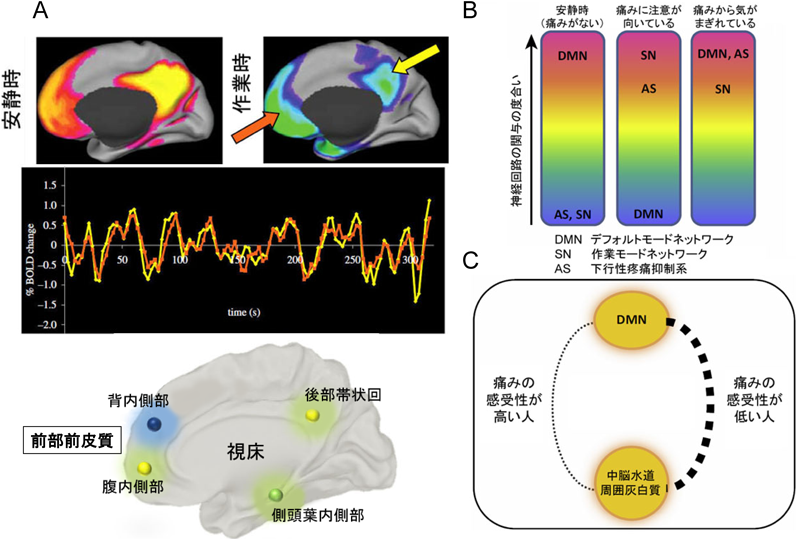
(A) DMNの部位とfMRIの活動変化の一致.(B)痛みの状態によりDMN,痛みのネットワーク(SN),下行性疼痛抑制系(AS)の関与の度合いの変化を示す.(C) DMNと中脳水道周囲灰白質との連結度合いと痛みの感受性.痛みに敏感な人はその連結が弱く,痛みにがまん強い人はその連結が強い.
最近,痛みに注意が向いているか,いないかに関わる三つのネットワークの関係が提案された.一つ目がデフォルトモードネットワーク,二つ目が痛みに意識がいっているときの痛みのネットワーク,三つ目が下行性疼痛抑制系である.痛みがない安静時には,デフォルトモードネットワークが強く関与し,図8Aのように赤くなる.痛みに注意が向いているときは,痛みのネットワークが活性化して,下行性疼痛抑制系とデフォルトモードネットワークの結びつきが弱くなっている.痛い状態はあっても,痛みから気がまぎれているときは,下行性疼痛抑制系が強く働き,デフォルトモードネットワークとの結びつきが強まっている(図8B).このことから,最近,痛みの感受性の違いは下行性疼痛抑制系とデフォルトモードネットワークの連結の強さの違いという考え方が提唱されている(図8C)49).痛みに対する感受性の高い人,いわゆる痛がり,はこの連結が弱く,感受性が低い人はこの連結が強いことになる.痛みの感受性が上がっている患者の痛みには,下行性疼痛抑制系に作用する薬が処方され,慢性腰痛のように器質的な疾患がない場合には,各個人の行動変容による痛みの治療,認知行動療法が取り入れられ始めている.
生化学の研究は,動物の病態・生理機能の解明から,タンパク質精製,遺伝子のクローニングによる機能分子の同定の時代を経て,動物個体,中でもヒトに対象が移っている.現在脚光を浴びているデフォルトモードネットワークを提唱したM.E. Raichleの総説48)の中に引用されている「ヒトは長寿で高齢社会を迎えているが,生まれたときにできたニューロンの大部分は,分裂することもなく取り替えられることもなく何10年生き続け,高次脳機能を担っている.イオンチャネルタンパク質,シナプス上の受容体や神経伝達に関わる機能分子は対照的に,早いもので数分,遅いものでも数週間で整備され,取り替えられている.どのニューロンも細胞内の分子的,生化学的な仕組みを利用してパーツを取り替え再構築している.このことにより脳は成長や学習に伴う可塑的な変化を可能にしているが,一方で神経機能を安定して維持するかという問題を抱えている.」50)は至言であり,これまで述べた痛みの発生・維持にも当てはまる.
本稿では,痛みの最新のメカニズムをできるだけ平易にわかりやすく記載したが,紙面の制約上,筆者が行った研究を含め痛みに関連するタンパク質分子の特定とそのアミノ酸配列に隠された機能に重点を置いた.私が40年以上医化学(生化学)を専攻してよかったと思うことは,生命の神秘の「パーツ」が次々と解き明かされた歴史的発見の時代にワクワク感を持って研究できたことである.「神経機能を安定して維持する」ことは個人のアイデンティティの基盤であり,その障害は認知症など高齢社会を迎えた日本の課題であるが,痛みでは不安や恐怖などの過去の負の記憶を消せないことが問題である.ヒトの脳科学の時代,本稿で十分記載できなかった痛みの情動認知面の理解が進み,痛みが克服される日がくることを期待している.
1) Norrsell, U., Finger, S., & Lajonchere, C. (1999) Brain Res. Bull., 48, 457–465.
2) Janscó, G., Kiraly, E., & Janscó-Gábor, A. (1977) Nature, 270, 741–743.
3) Caterina, M.J., Schumacher, M.A., Tominaga, M., Rosen, T.A., Levine, J.D., & Julius, D. (1997) Nature, 389, 816–824.
4) Tominaga, M., Caterina, M.J., Malmberg, A.B., Rosen, T.A., Gilbert, H., Skinner, K., Raumann, B.E., Basbaum, A.I., & Julius, D. (1998) Neuron, 21, 531–543.
5) Caterina, M.J., Rosen, T.A., Tominaga, M., Brake, A.J., & Julius, D. (1999) Nature, 398, 436–441.
6) McKemy, D.D., Neuhausser, W.M., & Julius, D. (2002) Nature, 416, 52–58.
7) Story, G.M., Peier, A.M., Reeve, A.J., Eid, S.R., Mosbacher, J., Hricik, T.R., Earley, T.J., Hergarden, A.C., Andersson, D.A., Hwang, S.W., McIntyre, P., Jegla, T., Bevan, S., & Patapoutian, A. (2003) Cell, 112, 819–829.
8) Coste, B., Mathur, J., Schmidt, M., Earley, T.J., Ranade, S., Petrus, M.J., Dubin, A.E., & Patapoutian, A. (2011) Science, 330, 55–60.
9) Maksimovic, S., Nakatani, M., Baba, Y., Nelson, A.M., Marshall, K.L., Wellnitz, S.A., Firozi, P., Woo, S.H., Ranade, S., Patapoutian, A., & Lumpkin, E.A. (2014) Nature, 509, 617–621.
10) Ikeda, R., Cha, M., Ling, J., Jia, Z., Coyle, D., & Gu, J.G. (2014) Cell, 157, 664–675.
11) Ranade, S.S., Woo, S.H., Dubin, A.E., Moshourab, R.A., Wetzel, C., Petrus, M., Mathur, J., Bégay, V., Coste, B., Mainquist, J., Wilson, A.J., Francisco, A.G., Reddy, K., Qiu, Z., Wood, J.N., Lewin, G.R., & Patapoutian, A. (2014) Nature, 516, 121–125.
12) Woo, S.H., Lukacs, V., de Nooij, J.C., Zaytseva, D., Criddle, C.R., Francisco, A., Jessell, T.M., Wilkinson, K.A., & Patapoutian, A. (2015) Nat. Neurosci., 18, 1756–1762.
13) McMillin, M.J., Beck, A.E., Chong, J.X., Shively, K.M., Buckingham, K.J., Gildersleeve, H.I., Aracena, M.I., Aylsworth, A.S., Bitoun, P., Carey, J.C., Clericuzio, C.L., Crow, Y.J., Curry, C.J., Devriendt, K., Everman, D.B., Fryer, A., Gibson, K., Giovannucci Uzielli, M.L., Graham, J.M. Jr., Hall, J.G., Hecht, J.T., Heidenreich, R.A., Hurst, J.A., Irani, S., Krapels, I.P., Leroy, J.G., Mowat, D., Plant, G.T., Robertson, S.P., Schorry, E.K., Scott, R.H., Seaver, L.H., Sherr, E., Splitt, M., Stewart, H., Stumpel, C., Temel, S.G., Weaver, D.D., Whiteford, M., Williams, M.S., Tabor, H.K., Smith, J.D., Shendure, J., Nickerson, D.A., & Bamshad, M.J.; University of Washington Center for Mendelian Genomics. (2014) Am. J. Hum. Genet., 94, 734–744.
14) Noda, M., Shimizu, S., Tanabe, T., Ikeda, T., Takahashi, H., Nakayama, H., Kanaoka, Y., Minamino, N., Kangawa, K., Matsuo, H., Raftery, M.A., Hiorse, T., Inayama, S., Hayashida, H., Miyata, T., & Numa, S. (1984) Nature, 312, 121–127.
15) Noda, M., Ikeda, T., Kayano, T., Suzuki, H., Takeshima, H., Kurasaki, M., Takahashi, H., & Numa, S. (1986) Nature, 320, 188–192.
16) Noda, M., Ikeda, T., Suzuki, H., Takeshima, H., Takahashi, T., Kuno, M., & Numa, S. (1986) Nature, 322, 826–828.
17) Cox, J.J., Reimann, F., Nicholas, A.K., Thornton, G., Roberts, E., Springell, K., Karbani, G., Jafri, H., Mannan, J., Raashid, Y., Al-Gazali, L., Hamamy, H., Valente, E.M., Gorman, S., Williams, R., McHale, D.P., Wood, J.N., Gribble, F.M., & Woods, C.G. (2006) Nature, 444, 894–898.
18) Yang, Y., Wang, Y., Li, S., Xu, Z., Li, H., Ma, L., Fan, J., Bu, D., Liu, B., Fan, Z., Wu, G., Jin, J., Ding, B., Zhu, X., & Shen, Y. (2004) J. Med. Genet., 41, 171–174.
19) Numazaki, M., Tominaga, T., Toyooka, H., & Tominaga, M. (2002) J. Biol. Chem., 277, 13375–13378.
20) Fitzgerald, E.M., Okuse, K., Wood, J.N., Dolphin, A.C., & Moss, S.J. (1999) J. Physiol., 516, 433–446.
21) Minami, T., Uda, R., Horiguchi, S., Ito, S., Hyodo, M., & Hayaishi, O. (1992) Pain, 50, 223–229.
22) Minami, T., Uda, R., Horiguchi, S., Ito, S., Hyodo, M., & Hayaishi, O. (1994) Pain, 57, 217–223.
23) Woolf, C.J., Shortland, P., & Coggeshall, R.E. (1992) Nature, 355, 75–78.
24) Ji, R.R., Kohno, T., Moore, K.A., & Woolf, C.J. (2003) Trends Neurosci., 26, 696–706.
25) Abe, T., Matsumura, S., Katano, T., Mabuchi, T., Takagi, K., Xu, L., Yamamoto, A., Hattori, K., Yagi, T., Watanabe, M., Nakazawa, T., Yamamoto, T., Mishina, M., Nakai, Y., & Ito, S. (2005) Eur. J. Neurosci., 22, 1445–1454.
26) Nakazawa, T., Komai, S., Tezuka, T., Hisatsune, C., Umemori, H., Semba, K., Mishina, M., Manabe, T., & Yamamoto, T. (2001) J. Biol. Chem., 276, 693–699.
27) Matsumura, S., Kunori, S., Mabuchi, T., Katano, T., Nakazawa, T., Abe, T., Watanabe, M., Yamamoto, T., Okuda-Ashitaka, E., & Ito, S. (2010) Eur. J. Neurosci., 32, 798–810.
28) Husi, H. & Grant, S.G. (2001) Trends Neurosci., 24, 259–266.
29) Mabuchi, T., Shintani, N., Matsumura, S., Okuda-Ashitaka, E., Hashimoto, H., Muratani, T., Minami, T., Baba, A., & Ito, S. (2004) J. Neurosci., 24, 7283–7291.
30) Ohnishi, T., Okuda-Ashitaka, E., Matsumura, S., Katano, T., Nishizawa, M., & Ito, S. (2008) J. Neurochem., 105, 2271–2285.
31) Roche, K.W., Standley, S., McCallum, J., Dune Ly, C., Ehlers, M.D., & Wenthold, R.J. (2001) Nat. Neurosci., 4, 794–802.
32) Kunori, S., Matsumura, S., Okuda-Ashitaka, E., Katano, T., Audoly, L.P., Urade, Y., & Ito, S. (2011) Glia, 59, 208–218.
33) Nishihara, I., Minami, T., Watanabe, Y., Ito, S., & Hayaishi, O. (1995) Neurosci. Lett., 196, 57–60.
34) Watabe, A., Sugimoto, Y., Honda, A., Irie, A., Namba, T., Negishi, M., Ito, S., Narumiya, S., & Ichikawa, A. (1993) J. Biol. Chem., 268, 20175–20178.
35) Tatsumi, S., Mabuchi, T., Katano, T., Matsumura, S., Abe, T., Hidaka, H., Suzuki, M., Sasaki, Y., Minami, T., & Ito, S. (2005) Neuroscience, 131, 491–498.
36) Okuda-Ashitaka, E., Minami, T., Matsumura, S., Takeshima, H., Reinscheid, R.K., Civelli, O., & Ito, S. (2006) Eur. J. Neurosci., 23, 995–1004.
37) Ahmadi, S., Lippross, S., Neuhuber, W.L., & Zeilhofer, H.U. (2002) Nat. Neurosci., 5, 34–40.
38) Inada, Y., Morimoto, S., Moroi, K., Endo, K., & Nakamura, T. (2005) Pain, 117, 251–258.
39) Unezaki, S., Yoshii, S., Mabuchi, T., Saito, A., & Ito, S. (2009) J. Neurosci. Methods, 178, 308–315.
40) Noda, M. & Sakuta, H. (2013) Trends Neurosci., 36, 661–673.
41) Unezaki, S., Katano, T., Hiyama, T.Y., Tu, N.H., Yoshii, S., Noda, M., & Ito, S. (2014) Eur. J. Neurosci., 39, 720–729.
42) Shimizu, H., Watanabe, E., Hiyama, T.Y., Nagakura, A., Fujikawa, A., Okado, H., Yanagawa, Y., Obata, K., & Noda, M. (2007) Neuron, 54, 59–72.
43) Tu, N.H., Katano, T., Matsumura, S., Funatsu, N., Pham, V.M., Fujisawa, J., & Ito, S. (2017) Eur. J. Neurosci., 46, 2096–2107.
44) Matsumura, S., Taniguchi, W., Nishida, K., Nakatsuka, T., & Ito, S. (2015) Eur. J. Neurosci., 41, 989–997.
45) Nakanishi, S., Inoue, A., Kita, T., Nakamura, M., Chang, A.C.Y., Cohen, S.N., & Numa, S. (1979) Nature, 278, 423–427.
46) Pavlov, I.P. 著,川村 浩訳(1975)大脳半球の働きについて—条件反射学—(上),pp. 54–56,岩波書店.
47) Roy, M., Shohamy, D., Daw, N., Jepma, M., Wimmer, G.E., & Wager, T.D. (2014) Nat. Neurosci., 17, 1607–1612.
48) Raichle, M.E. (2015) Phil.Trans. R. Soc. B, 370, 20140172.
49) Kucyi, A. & Davis, K.D. (2015) Trends Neurosci., 38, 86–95.
50) Marder, E. & Goaillard, J.M. (2006) Nat. Rev. Neurosci., 7, 563–574.

関西医科大学医化学講座教授.医学博士.
1950年大阪府に生る.76年京都大学医学部卒業.81年同大学院医学研究科修了.NIH(Fogartyフェロー),新技術開発事業団,大阪バイオサイエンス研究所を経て,94年より現職,2007年より同副学長.
研究テーマと抱負PARPのタンパク質精製,がん遺伝子産物Srcのリン酸化機構,ホルモン受容体と情報伝達機構,痛みの研究,特に神経障害性疼痛の発症・維持機構,に従事し,これからはヒトにおける痛みの研究に取り組みたいと考えている.
ウェブサイトhttp://www3.kmu.ac.jp/medchem/
趣味読書,庭いじり,将棋,ハイキング.

This page was created on 2017-11-08T13:15:38.317+09:00
This page was last modified on 2017-12-20T10:22:24.344+09:00
このサイトは(株)国際文献社によって運用されています。