細胞質における膜タンパク質の品質管理機構Quality control system of cytosolic membrane protein
千葉大学大学院理学研究院Graduate School of Science, Chiba University ◇ 〒263–8522 千葉市稲毛区弥生町1–33 ◇ 1–33 Yayoi, Inage-ku, Chiba 263–8522, Japan
発行日:2017年12月25日Published: December 25, 2017
© 2017 公益社団法人日本生化学会© 2017 The Japanese Biochemical Society
アミノ酸の性質を大きく分類すると親水性もしくは疎水性のものに分けられる.生体膜に挿入される膜貫通ドメインは約20個の疎水性アミノ酸配列からなり,さまざまなタンパク質ドメインの中でも最も高疎水性である.膜タンパク質は生体膜に挿入され機能するタンパク質であるが,膜貫通ドメインが細胞質へ露出すると,非特異的な疎水性相互作用を容易に引き起こし,凝集体形成の原因となるため,厳密に管理されなければならない.しかしタンパク質合成は細胞質で行われるため,膜貫通ドメインをいつどのようにして膜へ安全に挿入するか問題となる.特にテイルアンカー型膜タンパク質やミトコンドリア膜タンパク質は翻訳終了後に膜へと輸送されるため,膜貫通ドメインが細胞質に露出する危険性が高い.近年の研究から膜タンパク質の挿入経路だけでなく,分解経路についての分子メカニズムが明らかとなってきた.本稿では,膜挿入に失敗した小胞体とミトコンドリアの膜タンパク質の分解経路を中心に,膜タンパク質品質管理経路について概説する.
3種類の細胞小器官が膜タンパク質を直接膜挿入する機能を持つ.小胞体膜,ミトコンドリア膜,ペルオキシソーム膜である.小胞体は翻訳と共役した膜挿入経路と翻訳後に膜挿入される二つの挿入経路を持つ.I型,II型または複数回膜貫通型膜タンパク質は翻訳と共役して小胞体膜に挿入される.翻訳共役型膜挿入経路では,リボソームトンネルから翻訳途中のタンパク質の膜貫通ドメインが現れるとSRP(signal recognition particle)によって認識され,小胞体膜へターゲティングされ,翻訳しながら膜挿入される1)(図1).膜貫通ドメインがほとんど細胞質へ露出することなくリボソームから小胞体膜へと移行できることが翻訳共役型膜挿入経路の大きな利点である.一方,テイルアンカー型膜タンパク質は翻訳共役型膜挿入経路を利用できない.なぜならリボソーム内のトンネルはポリペプチド40残基分であり,膜貫通ドメインをC末端に持つテイルアンカー型の膜タンパク質は,膜貫通ドメインがリボソームから露出する前に翻訳が終了するため,リボソームから細胞質に放出されてしまう2).このため,テイルアンカー型タンパク質は翻訳終了後に細胞質のシャペロンに保護されながら目的の膜へ輸送される.細胞質に放出されたテイルアンカー型膜タンパク質の膜貫通ドメインはコシャペロンのSGTA(small glutamine-rich tetratricopeptide repeat-containing protein alpha)によって直接認識され,ATPase活性を持つTRC40(transmembrane domain recognition complex 40)に引き渡され,小胞体膜上の受容体であるGET1/2複合体を経由して,膜へ挿入される1).
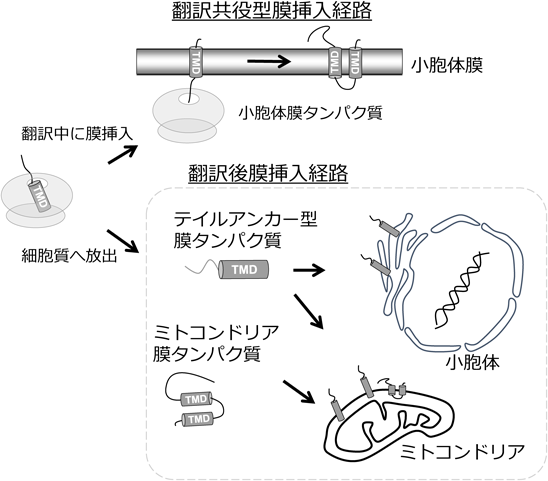
多くの小胞体膜タンパク質は翻訳中にリボソームが小胞体膜上に移行し,翻訳されながら膜挿入される翻訳共役型膜挿入経路をとる.一方で,テイルアンカー型膜タンパク質やミトコンドリアの膜タンパク質は翻訳が終了し,細胞質へ放出された後に目的の膜へ移行する.TMD:膜貫通ドメイン(transmembrane domain).
ミトコンドリアの膜タンパク質のほとんどは細胞質で合成され3),翻訳終了後にミトコンドリアへ輸送される.リボソームから放出されたミトコンドリア膜タンパク質はHsp70などのシャペロンの働きによりミトコンドリアへ輸送されると考えられている.ミトコンドリア外膜にあるTom複合体がミトコンドリア膜タンパク質を認識し,ミトコンドリア膜への挿入を仲介している4).テイルアンカー型膜タンパク質やミトコンドリアの膜タンパク質は翻訳終了後に細胞質に放出されるため,膜貫通ドメインが細胞質に露出する危険がある.高疎水性のアミノ酸からなる膜貫通ドメインの露出は,すぐさま非特異的な相互作用を引き起こし,タンパク質凝集体形成を誘導する.そのため細胞質に露出した膜タンパク質は後述するように特別な機構により品質管理されている.
小胞体テイルアンカー型膜タンパク質はリボソームから放出後,膜貫通ドメインがSGTAとTRC40に保護されながら膜へ移行する5).しかしSGTAとTRC40だけではテイルアンカー型膜タンパク質の膜移行に不十分であった.そこで同定されたのがBag6-TRC35-Ubl4A複合体である6, 7).Bag6複合体はSGTAとTRC40に結合できること,Bag6を欠損するとテイルアンカー型膜タンパク質とTRC40の結合が減少すること,Bag6もしくはTRC40の欠損はテイルアンカー型膜タンパク質とSGTAの結合を増加させることから,SGTAがテイルアンカー型膜タンパク質を最初に認識し,Bag6複合体上でSGTAからTRC40へ基質の受け渡しがされると考えられている(図2).興味深いことにBag6はリボソームと結合することが報告されており,リボソーム上で放出されたテイルアンカー型膜タンパク質は,リボソームと結合したBag6複合体のSGTAにすぐさま認識されているのかもしれない8).
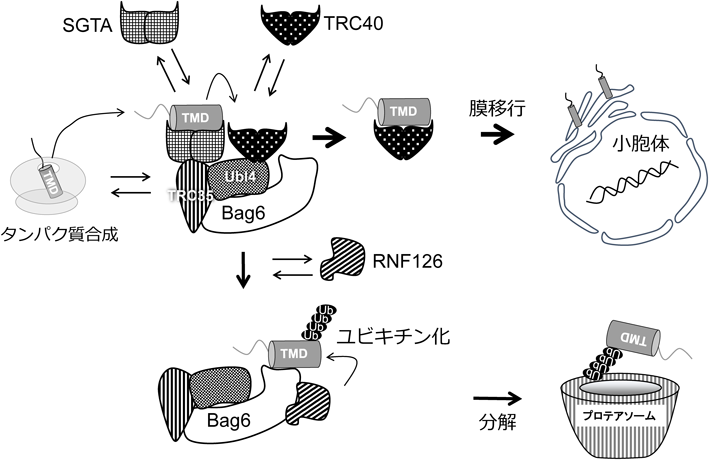
小胞体のテイルアンカー型膜タンパク質は翻訳終了後にBag6複合体上のSGTAに結合する.TRC40を介して小胞体膜上へ膜タンパク質が輸送され膜挿入される.小胞体膜挿入経路に障害があると,膜タンパク質がBag6複合体上に長くとどまるため,時間依存的にユビキチンリガーゼである126がBag6複合体にリクルートされ,膜タンパク質をユビキチン化し,プロテアソームによる膜タンパク質の分解を誘導する.
小胞体ストレスや,膜タンパク質の過剰産生,膜移行システムの変異などにより,膜タンパク質の高疎水性ドメインが細胞質に露出することが知られている9–11).Bag6複合体は上記で述べた膜タンパク質のターゲッティングだけでなく,細胞質に露出した膜タンパク質などを分解へ導く品質管理経路としても働く.哺乳類培養細胞から免疫沈降を用いて精製したBag6複合体と基質タンパク質のin vitroユビキチン化アッセイを行うと,基質がユビキチン化されることが判明した.ユビキチン化にはBag6複合体に含まれるUbl4AとTRC35は必要なく,Bag6のN末端に存在するUBL(ubiquitin-like)ドメインが必要であることから,Bag6のUBLドメインにユビキチンリガーゼが結合することが予想された.そのためBag6のUBLドメインに結合するユビキチンリガーゼを質量分析により決定したところRNF126が同定された.RNF126を欠損した細胞から精製したBag6複合体はユビキチンリガーゼ活性を消失すること,高疎水性ドメインを持つ基質がBag6-RNF126複合体によりユビキチン化を受けることなどがin vitro, in vivoの実験から示された12).これらのことから細胞質に露出した小胞体膜タンパク質の高疎水性ドメインはSGTAに認識された後,小胞体膜へ移行できない場合に,Bag6を介してRNF126がリクルートされ,分解経路へ誘導されることがわかった.ではSGTAに結合した膜タンパク質はどのようにしてTRC40による膜への移行とBag6-RNF126による分解経路へ選別されるのだろうか? この問題を解くため,精製したそれぞれの因子と光架橋を用いたin vitro実験により結合速度などを詳細に調べたところ,SGTAに結合した高疎水性ドメインをもつタンパク質はTRC40が存在すると,速やかにSGTAからTRC40へ受け渡されるのに対して,TRC40が何らかのトラブルによりBag6複合体上に存在しない場合(TRC40がすでに他のタンパク質に占められているなど)や,TRC40に対して指向性の低い高疎水性ドメインを持つタンパク質は,TRC40に受け渡されない.その場合,20秒程度経過すると膜タンパク質は自発的にSGTAから遊離し,Bag6へと受け渡されRNF126依存的に分解されるという「時間依存的なタンパク質品質管理モデル」が提唱されている13).
上述したとおり,小胞体の膜貫通ドメインを保護するシャペロンに関してはよく理解が進んでいる14).しかし細胞質からミトコンドリア膜タンパク質を保護する因子は明らかとなっていなかった.ミトコンドリアは約1000~1500種類のタンパク質を持ち,そのほぼすべてのタンパク質は細胞質で合成され,大部分が膜貫通ドメインを含んでいる.これまでの研究からミトコンドリアの外膜と内膜のタンパク質透過装置に関わる機構は明らかとされてきた15).また一般的なシャペロンであるHsp70とHsp90はさまざまなミトコンドリア外膜タンパク質の前駆体と結合し,折りたたみ状態を調節しながら,輸送に関わることが示唆されている.だがこれらのシャペロンが持つ基質結合サイトは典型的な膜貫通ドメインと効率的に結合するためには小さすぎる.さらに細胞質に存在するミトコンドリア膜タンパク質前駆体の保護,ならびに輸送が失敗した場合に分解へと導く細胞質因子はまったくわかっていなかった.
そこで筆者らはミトコンドリアテイルアンカー型膜タンパク質Omp25の膜貫通ドメインをモデル基質としてミトコンドリア膜タンパク質の移行もしくは品質管理経路の解明にチャレンジした.培養細胞を用いた実験では,タンパク質発現後にすぐさま膜へと挿入されてしまうため,細胞質に存在する膜タンパク質を捕らえることが困難である.そこでウサギ網状赤血球ライセートを用いたin vitroタンパク質合成系を利用した.ウサギ網状赤血球ライセートは膜成分を含まないため,in vitroで合成されたOmp25は赤血球由来の細胞質タンパク質と結合すると予想された.実際にin vitro合成したOmp25は可溶性を保ち,60~80 kDaと推定されるタンパク質と複合体を形成していた.Omp25に結合するタンパク質を質量分析を用いて解析したところUBQLN1, UBQLN2, UBQLN4がOmp25結合タンパク質として同定された(以下UBQLNとする)16).UBQLNはUBLとUBAドメインによってそれぞれプロテアソームとユビキチンに結合することから,ユビキチン化タンパク質をプロテアソームへ輸送するシャトル分子として機能することが想定されている.筆者らはUBQLNのSti様ドメインが他の膜タンパク質結合ドメインと似た高メチオニン領域であることを見いだし,膜タンパク質のシャペロンとしての機能もあわせ持つと予想した.
小胞体膜タンパク質とミトコンドリア膜タンパク質はどのように識別されるのだろうか? 重要なことに小胞体膜タンパク質の膜貫通ドメインは高疎水性である一方で,ミトコンドリアの膜貫通ドメインは疎水性度が若干低い.よって膜タンパク質へのシャペロンの結合量を調べてみると,小胞体膜タンパク質であるVAMP2やSec61bにはSGTA, TRC40が効率的に結合し,ミトコンドリア膜タンパク質であるOmp25やTom5にはSGTAやTRC40よりもUBQLNが効率的に結合した.したがって疎水性度の違いにより,それぞれのシャペロンの嗜好性が異なると考えられる.
UBQLNタンパク質の生理的重要性を調べるために,CRISPR/Cas9法を用いて三つのUBQLNをノックアウトしたUBQLN KO細胞を作製した.Omp25を野生型の細胞に発現させると,Omp25は界面活性剤に可溶性の画分(ミトコンドリアに膜挿入されたOmp25)に検出されるが,UBQLN KO細胞にOmp25を発現させると,界面活性剤に不溶性の画分にもOmp25が多く検出された.テイルアンカー型膜タンパク質ではない2回膜貫通型のミトコンドリア内膜タンパク質ATP5G1を用いた場合でも,同様の結果を得た.重要なことに,ミトコンドリア脱分極剤によるミトコンドリアストレスを加えると,野生型の細胞においてもATP5G1の前駆体の増加が観察された.さらに蓄積したATP5G1前駆体にはUBQLNが結合していることもわかった.これらのことは,ストレスを受けたミトコンドリアはトランスロコンによるタンパク質の取り込みを減少させる.その結果ミトコンドリアへ輸送できなくなった膜タンパク質は細胞質にとどまるが,UBQLNのシャペロン活性により凝集体形成から保護され,最終的に分解へ誘導されることが明らかとなった.
UBQLNがどのようにして膜タンパク質を分解へと導くのか調べるために,精製したUBQLN-Omp25複合体とユビキチン,E1, E2酵素を混合し,in vitroユビキチン化アッセイを行った.すると野生型のUBQLN複合体はOmp25をユビキチン化したが,UBQLNΔUBA(ubiquitin-associatingドメイン)変異体ではOmp25のユビキチン化がみられなかった.
さらに野生型UBQLNとOmp25の複合体を用いてミトコンドリアへのトランスロケーションアッセイを行うと,ユビキチン化されたOmp25はミトコンドリアへ移行されないが,UBQLNΔUBA変異体ではユビキチン化させたOmp25でもミトコンドリアへの移行が観察された.よってユビキチンリガーゼがUBQLNのUBAに結合し,基質をユビキチン化する.ユビキチン化された基質はUBAドメインに強く結合する.つまり不良膜タンパク質基質がUBQLNから離れミトコンドリアへ間違って移行することを抑制している.
UBQLNのUBLドメインはプロテアソームと結合するが,興味深いことにUBAドメインを欠損したUBQLN変異体とプロテアソームの結合は増加する.よってUBQLNは通常状態ではUBAとUBLドメインが分子内結合しており,ユビキチン化基質がUBAドメインに結合すると,フリーとなったUBLドメインが速やかにプロテアソームに結合すると考えられる.
以上の結果から細胞質に蓄積するミトコンドリア膜タンパク質の品質管理経路が浮かび上がった(図3).①細胞質に放出されたミトコンドリア膜タンパク質は,未解明の経路によりミトコンドリア膜へ移行する.膜タンパク質はUBQLNとも結合するが,結合は可逆的であり,ほとんどが膜へ移行される.②ミトコンドリアストレスなどによりミトコンドリア膜移行が停止すると細胞質のミトコンドリア膜タンパク質とUBQLNの結合が増加する.③UBQLNは通常UBAとUBLドメインが自己結合したクローズ構造を形成している.UBQLNとの結合が長くなると,自己解離したUBAドメインに未同定のユビキチンリガーゼがリクルートされ,基質のユビキチン化を行う.④ユビキチン化された膜タンパク質はUBAドメインと高親和性の結合をするため,膜への移行に戻ることはできなくなる.⑤フリーとなったUBLドメインはプロテアソームと結合し,ユビキチン化膜タンパク質を速やかに分解へと誘導する16).
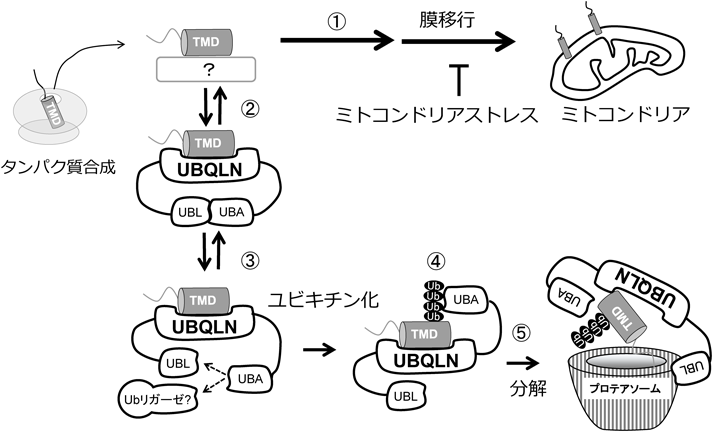
ミトコンドリアがストレスを受けると膜タンパク質の移行が抑制される.その結果,細胞質に増加したミトコンドリア膜タンパク質はUBQLNに結合し保護される.結合する時間が長くなると,UBQLNに未知のユビキチンリガーゼがリクルートされ,膜タンパク質をユビキチン化し,UBQLNのUBLドメインがプロテアソームをリクルートすることで,速やかな不要・不良膜タンパク質の分解を誘導する.
このようにUBQLNは細胞質のミトコンドリア膜タンパク質の認識から保護・分解まで一手に担うことで,いずれのステップにおいても膜貫通ドメインの露出を防ぐことができる新しいタイプの膜タンパク質品質管理経路である.
今回の発見から小胞体膜タンパク質だけでなく,ミトコンドリアの膜タンパク質も厳密に品質管理されていることが明らかとなった.実際にUBQLNファミリーの一つであるUBQLN2は筋萎縮性側索硬化症(ALS)の原因遺伝子として同定されていることから,膜タンパク質の細胞質での蓄積が神経疾患と関わることを示唆している17).一方で小胞体膜タンパク質の挿入経路に関わる因子がすべて同定されているにもかかわらず,いまだにミトコンドリア膜タンパク質の挿入経路に必須の因子は同定されていない.今後のさらなる研究からミトコンドリア膜タンパク質のターゲッティングの分子機構が解明されることが期待される.
1) Hegde, R.S. & Keenan, R.J. (2011) Nat. Rev. Mol. Cell Biol., 12, 787–798.
2) Kutay, U., Hartmann, E., & Rapoport, T.A. (1993) Trends Cell Biol., 3, 72–75.
3) Neupert, W. (1997) Annu. Rev. Biochem., 66, 863–917.
4) Young, J.C., Hoogenraad, N.J., & Hartl, F.U. (2003) Cell, 112, 41–50.
5) Shao, S. & Hegde, R.S. (2011) Annu. Rev. Cell Dev. Biol., 27, 25–56.
6) Hessa, T., Sharma, A., Mariappan, M., Eshleman, H.D., Gutierrez, E., & Hegde, R.S. (2011) Nature, 475, 394–397.
7) Mariappan, M., Mateja, A., Dobosz, M., Bove, E., Hegde, R.S., & Keenan, R.J. (2011) Nature, 477, 61–66.
8) Mariappan, M., Li, X., Stefanovic, S., Sharma, A., Mateja, A., Keenan, R.J., & Hegde, R.S. (2010) Nature, 466, 1120–1124.
9) Kang, S.W., Rane, N.S., Kim, S.J., Garrison, J.L., Taunton, J., & Hegde, R.S. (2006) Cell, 127, 999–1013.
10) Orsi, A., Fioriti, L., Chiesa, R., & Sitia, R. (2006) J. Biol. Chem., 281, 30431–30438.
11) Zimmermann, R., Müller, L., & Wullich, B. (2006) Trends Mol. Med., 12, 567–573.
12) Rodrigo-Brenni, M.C., Gutierrez, E., & Hegde, R.S. (2014) Mol. Cell, 55, 227–237.
13) Shao, S., Rodrigo-Brenni, M.C., Kivlen, M.H., & Hegde, R.S. (2017) Science, 355, 298–302.
14) Casson, J., McKenna, M., & High, S. (2016) Biochem. Soc. Trans., 44, 796–801.
15) Chacinska, A., Koehler, C.M., Milenkovic, D., Lithgow, T., & Pfanner, N. (2009) Cell, 138, 628–644.
16) Itakura, E., Zavodszky, E., Shao, S., Wohlever, M.L., Keenan, R.J., & Hegde, R.S. (2016) Mol. Cell, 63, 21–33.
17) Deng, H.X., Chen, W., Hong, S.T., Boycott, K.M., Gorrie, G.H., Siddique, N., Yang, Y., Fecto, F., Shi, Y., Zhai, H., Jiang, H., Hirano, M., Rampersaud, E., Jansen, G.H., Donkervoort, S., Bigio, E.H., Brooks, B.R., Ajroud, K., Sufit, R.L., Haines, J.L., Mugnaini, E., Pericak-Vance, M.A., & Siddique, T. (2011) Nature, 477, 211–215.

千葉大学大学院理学研究院助教.博士(理学).
2009年埼玉大学大学院理工学研究科博士課程修了,同年東京医科歯科大学大学院医歯学総合研究科研究員,12年英国MRC Laboratory of Molecular Biology研究員を経て,15年より千葉大学大学院助教.
研究テーマと抱負タンパク質の品質管理機構の研究.知りたいことを知るための努力を怠らないこと.
ウェブサイトhttp://life.s.chiba-u.jp/matsuura/HP/matsuura.html
趣味インドホシガメ(亀吾郎)の飼育と,カメを他人に勧めること.

This page was created on 2017-10-31T11:49:35.897+09:00
This page was last modified on 2017-12-20T10:27:14.55+09:00
このサイトは(株)国際文献社によって運用されています。