ペプチドミクスを活用した生理活性ペプチドの発見Peptidomics for bioactive peptide discovery
国立循環器病研究センターNational Cerebral and Cardiovascular Center ◇ 〒565–8565 大阪府吹田市藤白台5–7–1 ◇ 5–7–1 Fujishirodai, Suita, Osaka 565–8565, Japan
発行日:2017年12月25日Published: December 25, 2017
© 2017 公益社団法人日本生化学会© 2017 The Japanese Biochemical Society
ペプチドホルモンや神経ペプチドのような生理活性ペプチドは細胞間情報伝達分子の一種で,インスリンはその代表例である.生理活性ペプチドの新たな発見は基礎研究の分野にとどまらず,創薬をはじめとして各方面に波及効果をもたらす.
20世紀初頭のセクレチンの発見以来,生理活性ペプチドの大半は特定の生物活性や生化学的な応答を指標として発見されてきた.1990年代には,生理活性ペプチドの受容体の多くがGタンパク質と共役した7回膜貫通型受容体(GPCR)であることが明らかになった.オーファンGPCRのリガンド探索が進められ,日本からもオレキシンやグレリンなどが発見された.また,生理活性ペプチドの半数近くはそのC末端がアミド化修飾されていることや,無脊椎動物では,RFアミドと総称される生理活性ペプチド群が発見されるようになると,ゲノムからC末端アミド化ペプチドの生成部位が予測されるようになった.しかし,このようなアプローチで発見される新規ペプチドは近年減少している.
今世紀になり,アミノ酸配列決定の手段として質量分析が一般化した.我々は,ペプチドホルモン産生細胞が分泌するペプチドをそのままの分子型で一斉に分析すると,新規生理活性ペプチド発見の手がかりが得られるのではないかと考えた.得られた情報から候補ペプチドを予測し,機能解析を経ていくつかの新しい生理活性ペプチドを発見できた.本稿では,このようなペプチドミクスの考え方に基づいた生理活性ペプチド探索のアプローチとその実例について概説する.
ペプチドミクスは,細胞が産生するペプチド全体を解析する技術または研究分野を指す1, 2).ペプチドミクスで定義されるペプチドとは,翻訳されたタンパク質が内因性のプロテアーゼで切断されて生じるペプチドを指し,生理活性ペプチドもこの中に含まれる.一方,プロテオーム解析で使われる「ペプチド」は,基質特異性が明確な特定の酵素でin vitroで消化して得られた「断片ペプチド」を意味する.プロテオーム解析では,断片ペプチドの配列決定によってタンパク質を同定したと判断する.この点がペプチドミクスとの大きな違いである.
ゲノム情報が明らかな生物では,生物試料中に含まれる膨大な種類のペプチドの配列の同定は,液体クロマトグラフィー・タンデム質量分析法(LC-MS/MS)の技術で初めて可能となり,ペプチドミクス研究の進展に多大な貢献をしている.試料の分離には逆相系のカラムが主に利用され,試料中のペプチドはオンライン接続されたタンデム質量分析計に導かれる.多くの生理活性ペプチドは3000 Daを超えており,エレクトロスプレーイオン化法(ESI)が広範な質量範囲に対応できる.「断片」ペプチドに比較して,ペプチドミクスが扱う内在性のペプチドの配列決定はさまざまな制約により容易ではないため,ペプチドミクス研究は注目度が低いのは事実であるが,最新の質量分析計により現在は4桁に及ぶ異なる配列が一つの試料から同定可能である.本稿では定性的な解析での所見を述べるが,安定同位体標識を用いた定量的な解析法も海外から報告されている3).
ペプチドホルモンや神経ペプチドのような分泌性因子を質量分析で探索するためには,分泌ペプチドを豊富に含む試料が望ましい.我々は,分泌顆粒を持つ内分泌系の付着系細胞株を用い,数分間の脱分極刺激を与えて回収した培養上清を探索材料としている.これらの細胞は分泌顆粒の内容物を速やかに培地中に放出する.回収した培養上清をゲルろ過で分離し,ゲル電気泳動が不得手とする約1000から10,000 Daの画分をLC-MS/MS測定用の試料とする.同定ペプチドの95%以上はシグナルペプチドを持つ前駆体に由来していた4).分泌ペプチドを効率よく同定できるため,このような試料をsecretopeptidomeと呼んでいる(図1).
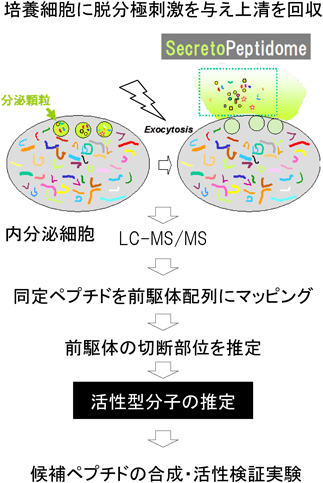
分泌顆粒を持つ内分泌細胞は脱分極刺激を与えると短時間のうちに顆粒内容物を細胞外に放出(exocytosis)する.培養細胞ではその培養上清を集めることで効率よく分泌ペプチドを回収できる.
同定されたペプチド群を前駆体に帰属させていくと,図2のようなマップが得られる.図はグレリン前駆体にマップされたペプチド群の一部を示しており,3番目のセリンがオクタン酸修飾された成熟型のグレリンが多数同定されている.同定ペプチド群がラダーを呈しているのは,プロセシング後にプロテアーゼで消化されていくためである.プロセシング情報が不明な前駆体タンパク質についても,同定ペプチドを前駆体に並べていくと,プロセシング部位が浮かび上がってくる5, 6).ペプチドホルモンや神経ペプチド前駆体は,塩基性アミノ酸対または,周辺アミノ酸に特定の傾向を持つ単一塩基性アミノ酸のC末端側で切断されることが経験的に知られているが,図からも明らかなようにそのような配列が常に切断されるわけではない.したがって,実在ペプチドを分析するペプチドミクスは,他の手法では得られない所見を提供している.我々は実際のプロセシング部位から新規の生理活性ペプチドを予測し,発見につなげることに世界で初めて成功した7).また,分泌タンパク質ばかりでなく,膜タンパク質の細胞外ドメインの切断部位なども浮かび上がってきた8).

マウス由来MGN3-1細胞のsecretopeptidomeを質量分析し,グレリン前駆体に帰属される配列をマッピングした.細い矩形が同定配列を示す.グレリンのオクタン酸修飾を黒で,プロペプチドのトレオニンのリン酸化を灰色で示す.グレリンは27および28アミノ酸の分子種が主要であり,そのことがこのマップからも確認され,シグナルペプチドの末端とグレリンのC末端のプロセシング部位(↓)が明瞭に浮かび上がっている.必ずしもすべての塩基性アミノ酸(白抜き文字)のC末端で切断されているわけではないことがわかる.文献6)より改変して記載.
分泌顆粒を持たない通常の培養細胞では,急速な分泌誘導は起こらず,回収までに数時間が必要となるので,細胞への影響について個別に対応が必要である.高等生物の場合,組織抽出物は細胞内タンパク質の分解ペプチドを多量に含んでいるので既知の生理活性ペプチドの同定も容易とはいえない.初代培養細胞は無血清培養に弱いため,分泌ペプチドの確保が難しい.
4桁に及ぶペプチド配列情報の中から生理活性ペプチドを発見するためには,候補ペプチドを選択し,合成ペプチドを用いた検証実験が必要である(図1).我々は種を超えて保存されている配列の中から,いくつかの指標を目安として選択している.本稿では,1)C末端アミド化,2)塩基性アミノ酸の存在割合,3)セカンドメッセンジャーとしての細胞内カルシウム上昇,のそれぞれを指標として候補ペプチドを選択し,機能解析を経て同定できた事例について述べる.
生理活性ペプチドの半数近くはC末端がアミド化されていることに着目し,新規のC末端アミド化ペプチドを同定する過程でNERP-1およびNERP-2は見いだされた7).カルシトニンやガストリン放出ペプチドなどの既知のC末端アミド化ペプチドを分泌するヒト甲状腺髄様がんのTT細胞株のsecretopeptidomeを回収し,LC-MS/MSによって,分泌タンパク質VGFの異なる領域から二つの新規C末端アミド化ペプチドが同定された(図3).VGFはニューロンや内分泌細胞で発現しており,1985年に神経栄養因子NGFによってPC12細胞で誘導される遺伝子としてクローニングされていた9).しかし,C末端アミド化ペプチド生成の報告はなかった.我々は,マウスの視床下部での生理的刺激によるVGF遺伝子発現の変化のデータから,水・電解質代謝への関与が示唆され,機能解析を進めたところ,この二つのペプチドが視索上核,室傍核の大細胞性神経分泌ニューロンに存在し,抗利尿ホルモン(AVP)の分泌を抑制することを明らかにし,NERP-1,-2と命名した.この作用はC末端アミド化構造に依存している.NERP-1は大細胞性神経分泌ニューロンとシナプスを作るグルタミン酸作動性ニューロンをシナプス前のレベルで抑制し,NERP-2はGABA作動性介在ニューロンを活性化し,シナプス前のレベルでグルタミン酸作動性ニューロンを抑制してAVP放出を抑制すると考えられる.
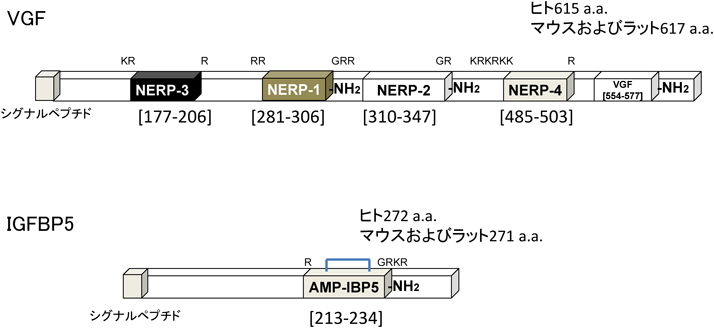
ペプチドのプロセシング部位に存在する塩基性アミノ酸(K, R)およびアミド化シグナル(GRR, GR, GRKR)を示す.VGFのC末端側のアミド化ペプチドは,マウス・ラットには存在しない.アミノ酸番号はヒトの配列に基づく.a.a.:アミノ酸.–NH2:C末端アミド化.
また,摂食中枢である視床下部外側野でVGF遺伝子の発現が絶食によって強く誘導された.脳室内投与されたNERP-2は摂食を亢進させる10).この作用はNERP-2のみで,かつアミド化依存性に認められる.オレキシンの中和抗体の脳室内投与,または,オレキシン遺伝子ノックアウトマウスではNERP-2の作用は消失することから,オレキシンを介していると考えられる.末梢投与されたNERP-2は,高血糖で誘導されるインスリン分泌を亢進させる11).同様の作用を示すグルカゴン様ペプチド-1(GLP-1)に比較すると活性は1/100程度であるが,NERP-2は,膵のβ細胞のみならず消化管の内分泌細胞にも発現していることから,GLP-1と同様にインクレチンとして作用している可能性も示唆される.
生理活性ペプチドの多くが塩基性アミノ酸を多く含むことに着目し,ソマトスタチンを産生するヒト膵内分泌腫瘍培養株QGP-1のsecretopeptidomeを探索してAMP-IBP5は見いだされた12).前駆体はinsulin-like growth factor-binding protein 5(IGFBP5)で,1組の分子内ジスルフィド結合で環状化した22アミノ酸からなるC末端アミド化ペプチドである.N末端は,ペプチドホルモンのプロセシング酵素の認識配列に合致していた.ペプチドの配列はヒト,マウス,ラット,ブタ,ウシで共通で,ラット脳・腸においても同じ分子型で存在する.強い抗菌活性を持ち,検討した8種の菌のうち6種に対して,ヒトの主要な抗菌ペプチドであるβ-defensin-2より強く,cathelicidinに匹敵する活性を示した.IGFBP-5から機能性のペプチドが生成することは初めて得られた所見である.抗体を用いた検討によりAMP-IBP5は脳,下垂体,消化管に多く存在することから,IGFBP5とプロセシング酵素類を共発現する神経細胞・内分泌細胞が産生していると考えられる.
前述のVGF前駆体から同定された新規の抗菌ペプチドである5).配列内部に連続する塩基性アミノ酸があり,通常の衝突誘起開裂法では同定されず,電子転移開裂法によって同定された.以上の四つのペプチドはこれまでのin silicoの検索では未同定のペプチドであった.
前述のTT細胞のsecretopeptidome解析で同定したペプチド群からプロセシング部位を推定し,種間で相同性の高いペプチドを選択した.カルシウム感受性発光タンパク質エクオリンのトランスジェニックマウスの組織片に候補ペプチドを添加して発光量の変化を評価するスクリーニングを実施した.VGF[177–206],[485–503]に相当するペプチドが下垂体,視床下部の組織片にカルシウム応答を誘導したので,これらをNERP-3およびNERP-4と命名した13).室傍核および視索上核のAVP陽性神経分泌ニューロンの中にNERP-3に反応する細胞があり,in vitroでNERP-3はラットの下垂体後葉からAVPの分泌を促す14).
生理活性ペプチドの新しい探索法について述べた.NERP-1,-2は質量分析を活用して生理活性ペプチドの発見に至った初の事例である.その後,視交叉上核を含むラットの脳切片を電気刺激し,その放出物の解析からグラニン前駆体SAAS由来で生物時計を変化させる活性ペプチドが海外より報告されている15).候補ペプチドの活性を簡便にスクリーニングする方法の開発が課題の一つである.secretopeptidomeの質量分析に端を発して,生理活性ペプチドの発見が促進されることが期待される.
本研究は高尾敏文教授(大阪大学),中里雅光教授,山崎基生博士(宮崎大学),塩田清二教授(星薬科大学),上田陽一教授(産業医科大学),十枝内厚次教授(至学館大学),尾崎司博士(山形大学),Luis Gonzalez博士(国立遺伝子工学センター,キューバ),里見佳典博士(武田薬品工業),土谷翔氏,松原真佐子氏,南野直人博士,寒川賢治博士(国立循環器病研究センター)のご協力で得られた成果であり,深く感謝いたします.
1) Sasaki, K., Sato, K., Akiyama, Y., Yanagihara, K., Oka, M., & Yamaguchi, K. (2002) Cancer Res., 62, 4894–4898.
2) Schrader, M., Schultz-Nappe, P., & Fricker, L.D. (2014) EuPA Open Proteom., 3, 171–182.
3) Che, F.Y. & Fricker, L.D. (2005) J. Mass Spectrom., 40, 238–249.
4) Sasaki, K., Satomi, Y., Takao, T., & Minamino, N. (2009) Mol. Cell. Proteomics, 8, 1638–1647.
5) Sasaki, K., Osaki, T., & Minamino, N. (2013) Mol. Cell. Proteomics, 12, 700–709.
6) Tsuchiya, T., Iwakura, H., Minamino, N., Kangawa, K., & Sasaki, K. (2017) Biochem. Biophys. Res. Commun., 490, 1142–1146.
7) Yamaguchi, H., Sasaki, K., Satomi, Y., Shimbara, T., Kageyama, H., Mondal, M.S., Toshinai, K., Date, Y., González, L.J., Shioda, S., Takao, T., Nakazato, M., & Minamino, N. (2007) J. Biol. Chem., 282, 26354–26360.
8) Tsuchiya, T., Osaki, T., Minamino, N., & Sasaki, K. (2015) J. Proteome Res., 14, 4921–4931.
9) Levi, A., Eldridge, J.D., & Paterson, B.M. (1985) Science, 229, 393–395.
10) Toshinai, K., Yamaguchi, H., Kageyama, H., Matsuo, T., Koshinaka, K., Sasaki, K., Shioda, S., Minamino, N., & Nakazato, M. (2010) Am. J. Physiol. Endocrinol. Metab., 299, E394–E401.
11) Moin, A.S., Yamaguchi, H., Rhee, M., Kim, J.W., Toshinai, K., Waise, T.M., Naznin, F., Matsuo, T., Sasaki, K., Minamino, N., Yoon, K.H., & Nakazato, M. (2012) Biochem. Biophys. Res. Commun., 428, 512–517.
12) Osaki, T., Sasaki, K., & Minamino, N. (2011) J. Proteome Res., 10, 1870–1880.
13) Sasaki, K., Takahashi, N., Satoh, M., Yamasaki, M., & Minamino, N. (2010) J. Proteome Res., 9, 5047–5052.
14) Fujihara, H., Sasaki, K., Mishiro-Sato, E., Ohbuchi, T., Dayanithi, G., Yamasaki, M., Ueta, Y., & Minamino, N. (2012) Endocrinology, 153, 1377–1386.
15) Hatcher, N.G., Atkins, N.Jr., Annangudi, S.P., Forbes, A.J., Kelleher, N.L., Gillette, M.U., & Sweedler, J.V. (2008) Proc. Natl. Acad. Sci. USA, 105, 12527–12532.

国立循環器病研究センター分子薬理部室長.博士(医学).
1990年東京大学医学部医学科卒業.97年国立がんセンター研究所細胞増殖因子研究部室長.2004年より現職.
研究テーマと抱負質量分析を活用した生理活性ペプチドの探索と機能解析.
趣味日本庭園鑑賞.クラシック音楽鑑賞.

This page was created on 2017-11-01T15:35:57.871+09:00
This page was last modified on 2017-12-20T10:31:11.195+09:00
このサイトは(株)国際文献社によって運用されています。