1) Wang, J., Alexander, P., Wu, L., Hammer, R., Cleaver, O., & McKnight, S.L. (2009) Science, 325, 435–439.
2) Shyh-Chang, N., Locasale, J.W., Lyssiotis, C.A., Zheng, Y., Teo, R.Y., Ratanasirintrawoot, S., Zhang, J., Onder, T., Unternaehrer, J.J., Zhu, H., Asara, J.M., Daley, G.Q., & Cantley, L.C. (2013) Science, 339, 222–226.
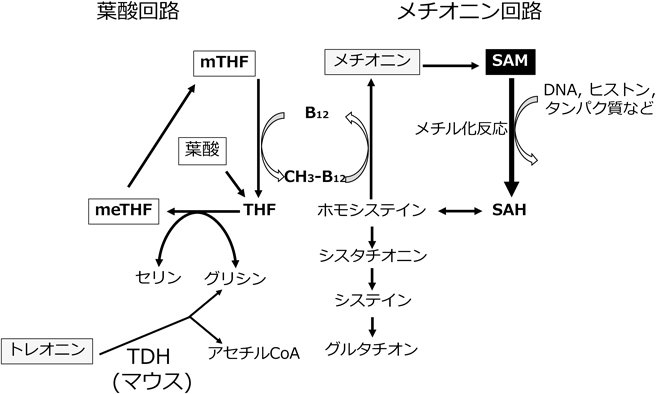
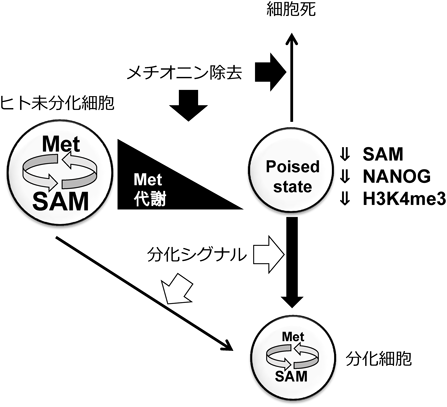
3) Shiraki, N., Shiraki, Y., Tsuyama, T., Obata, F., Miura, M., Nagae, G., Aburatani, H., Kume, K., Endo, F., & Kume, S. (2014) Cell Metab., 19, 780–794.
4) Mentch, S.J., Mehrmohamadi, M., Huang, L., Liu, X., Gupta, D., Mattocks, D., Gomez Padilla, P., Ables, G., Bamman, M.M., Thalacker-Mercer, A.E., Nichenametla, S.N., & Locasale, J.W. (2015) Cell Metab., 22, 861–873.
5) Li, S., Swanson, S.K., Gogol, M., Florens, L., Washburn, M.P., Workman, J.L., & Suganuma, T. (2015) Mol. Cell, 60, 408–421.
6) Matsuura, K., Kodama, F., Sugiyama, K., Shimizu, T., Hagiwara, N., & Okano, T. (2015) Tissue Eng. Part C Methods, 21, 330–338.
7) Saito, Y., Iwatsuki, K., Hanyu, H., Maruyama, N., Aihara, E., Tadaishi, M., Shimizu, M., & Kobayashi-Hattori, K. (2017) Biochem. Biophys. Res. Commun., 488, 171–176.
8) Sperber, H., Mathieu, J., Wang, Y., Ferreccio, A., Hesson, J., Xu, Z., Fischer, K.A., Devi, A., Detraux, D., Gu, H., Battle, S.L., Showalter, M., Valensisi, C., Bielas, J.H., Ericson, N.G., Margaretha, L., Robitaille, A.M., Margineantu, D., Fiehn, O., Hockenbery, D., Blau, C.A., Raftery, D., Margolin, A.A., Hawkins, R.D., Moon, R.T., Ware, C.B., & Ruohola-Baker, H. (2015) Nat. Cell Biol., 17, 1523–1535.
9) Tang, X., Keenan, M.M., Wu, J., Lin, C.A., Dubois, L., Thompson, J.W., Freedland, S.J., Murphy, S.K., & Chi, J.T. (2015) PLoS Genet., 11, e1005158.