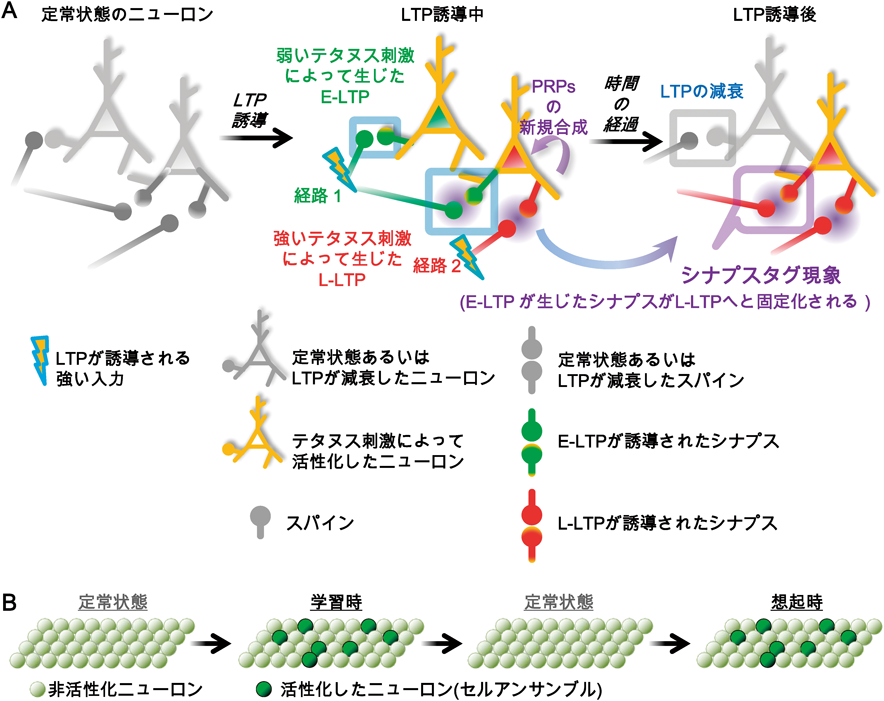
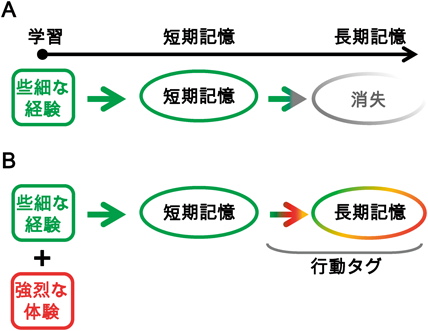
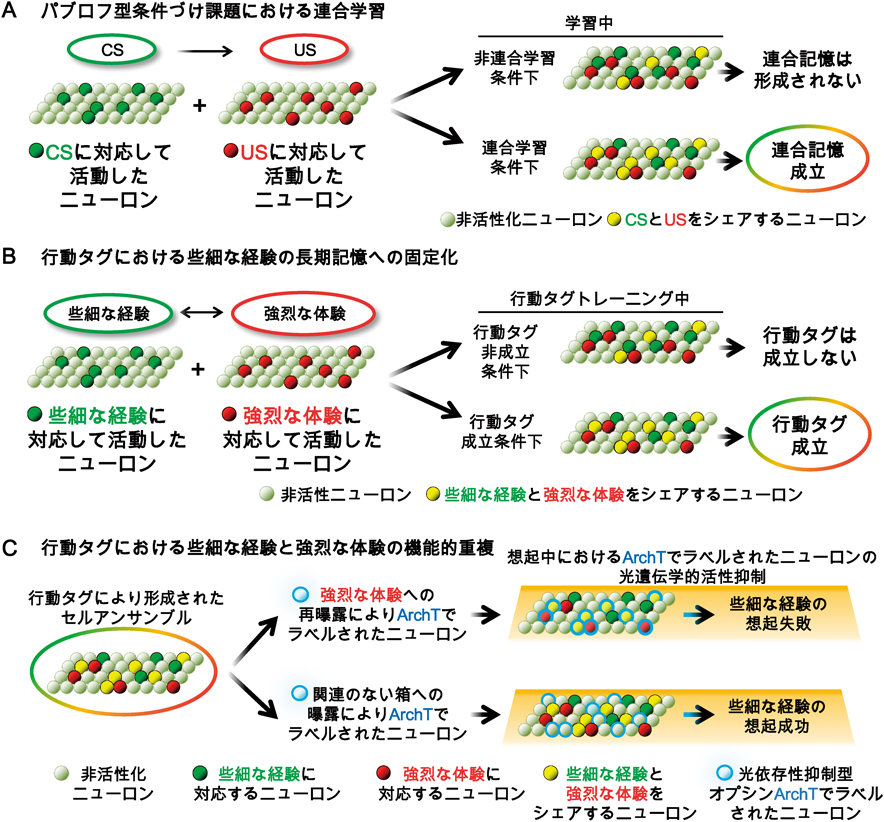
細胞タグ機構:行動タグのための記憶アロケーションの空間的重複Cellular tagging framework: The spatial overlapping of ensemble allocation for behavioral tagging
1 富山大学大学院医学薬学研究部(医学)生化学講座Department of Biochemistry, Graduate School of Medicine and Pharmaceutical Sciences, University of Toyama ◇ 〒930–0194 富山県富山市杉谷2630 ◇ 2630 Sugitani, Toyama 930–0194, Japan
2 JST・CRESTJST・CREST
発行日:2018年2月25日Published: February 25, 2018