シアリダーゼは,糖鎖から酸性糖であるシアル酸を遊離させる加水分解酵素である.シアル酸は主に糖鎖の末端を修飾しており,生理機能や疾患,ウイルス感染など幅広い生命現象に関わる.特に,シアル酸の基本構造であるノイラミン酸(neuraminic acid)の語源が神経(neuron)に由来することから象徴されるように,シアル酸は脳に豊富に含まれ,神経機能を発揮する上で欠かすことができない.したがって,シアリダーゼによるシアル酸脱離を介した糖鎖構造の制御もまた神経機能に大きな影響を与える.
哺乳動物のシアリダーゼには,細胞内局在や基質特異性,至適pHなどが異なるNeu1, Neu2, Neu3, Neu4の4種類のアイソザイムがあり,これらのシアリダーゼアイソザイムはすべて哺乳動物の脳に発現している1).最近では,外界からの刺激に応じてダイナミックに局在を変化させるなど,シアリダーゼの多彩な機能が明らかになりつつある2).本稿では筆者らの研究成果を中心に,記憶におけるシアリダーゼの役割について概説する.
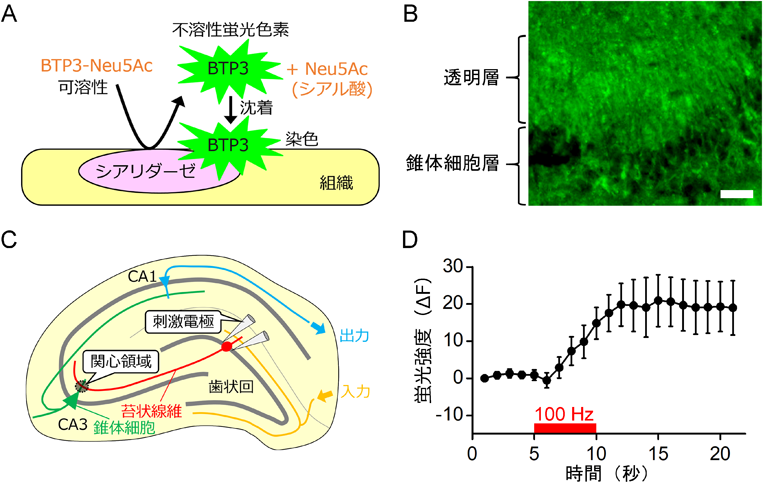
従来のシアリダーゼ活性測定用の人工基質では,動物シアリダーゼの酵素活性を組織上で高感度に可視化することは困難であった.筆者らは,シアリダーゼ活性の分布をイメージングできるプローブBTP-Neu5Acを開発した3).蛍光発色団の構造と励起/蛍光波長の違いにより,BTP2-Neu5Ac, BTP3-Neu5Ac, BTP4-Neu5Acの3種を作製した.BTP3-Neu5Acを例にとって染色原理を図1Aに示す.BTP3-Neu5Acは水に可溶であり無蛍光であるが,シアリダーゼによって加水分解を受けると,強い蛍光を持ったBTP3(励起波長374 nm,蛍光波長542 nm)を遊離する.BTP3は水に不溶であるため,組織に沈着することによって染色することができる.BTP4-Neu5Acは組織上で最も視認性がよいが,視認性と合成の簡便性のバランスから主にBTP3-Neu5Acを利用している.BTP3-Neu5Acは平成29年10月現在,和光純薬工業株式会社から入手できる.なお,BTP(2-benzothiazol-2-yl-phenol)は有機EL材料として研究されていた過去がある.長いストークスシフトを有するため励起光と蛍光の分離は比較的容易であるが,蛍光顕微鏡に標準で内蔵されているフィルターセットでは観察できない場合があるので注意が必要である.BTP-Neu5Acは現在,がんやウイルスの検出などに利用範囲を広げている.
BTP3-Neu5Acを利用して記憶をつかさどる海馬を染色すると,海馬の主要な興奮性神経である苔状線維の終末領域(CA3透明層)に比較的強い蛍光が観察される(図1B)4).海馬切片のシアリダーゼ活性をBTP3-Neu5Acでモニターした状態で苔状線維にLTP誘導性の高頻度電気刺激を与えると,刺激開始から1~2秒程度遅れて透明層の蛍光強度が増加する(図1C, D).このように,シアリダーゼ活性は神経発火と連動して迅速に酵素活性を変化させる.
シアリダーゼ活性の増加は,高頻度刺激以外にも脱分極刺激やフォルスコリンによる薬物性LTP誘導刺激,脳由来神経栄養因子(BDNF),NMDA受容体やカイニン酸受容体などのグルタミン酸受容体活性化によっても誘導される4).神経活動と連動してシアリダーゼ活性が増加する詳細な機構は不明であるが,脱分極刺激によってインキュベート液中のシアリダーゼ活性が変化しないことや各種シアリダーゼアイソザイムのmRNA量が増加しないことを考慮すると,シアリダーゼの細胞外への分泌や生合成量の変化によるものではなく,細胞内の局在変化や活性化因子などの影響により細胞膜表面の酵素活性が変化しているものと推定される.また,海馬の薄切切片を利用したシアリダーゼ活性の高解像度イメージングにより,苔状線維の投射先であるCA3錐体神経細胞の細胞膜において,脱分極刺激後にシアリダーゼ活性が増加していることが見いだされた.初代培養グリア細胞のシアリダーゼ活性もまた,グルタミン酸を添加することによって増加する.シアリダーゼ活性は,神経細胞のみでなくグリア細胞でも神経活動と連動して増加すると考えられる.
神経活動と連動した糖鎖からのシアル酸脱離について,レクチン染色を利用して検証した4).高濃度カリウムによって脱分極させた後の海馬切片では,シアル酸を認識するMAAレクチン[認識部位:Neu5Acα2-3Gal-β(1-3)-GalNAc]の結合が低下した.逆に,シアル酸脱離後に露出するガラクトース末端を認識するPNAレクチン[認識部位:Gal-β(1-3)-GalNAc]の結合は,脱分極後に増加した.このように,神経発火によるシアリダーゼ活性の増加は,糖鎖からシアル酸を脱離させるのに十分であることが示唆される.
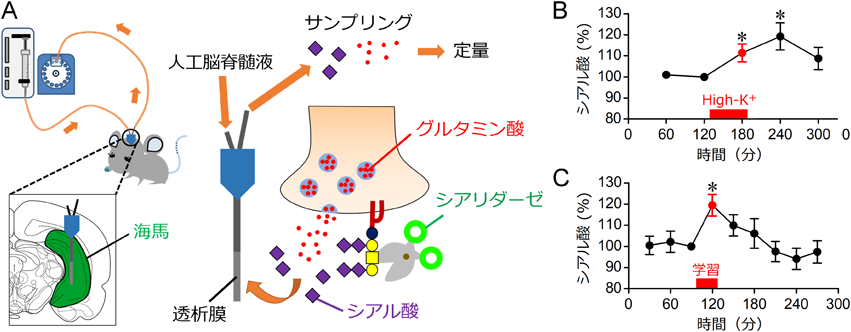
次に,in vivo(インビボ)マイクロダイアリシス法を利用して神経活動と連動したシアル酸脱離をインビボ条件下で検出することを試みた4).インビボマイクロダイアリシス法は微小透析膜を介して組織から低分子を直接回収することができる(図2A).高濃度カリウムでラットの海馬神経を脱分極させると,海馬の細胞外液から回収される遊離シアル酸量が増加した(図2B).海馬における遊離シアル酸量の増加は,恐怖条件づけ文脈学習時にも検出された(図2C).記憶形成時の神経活動と連動して,シアル酸が脱離することが見いだされた.
シアリダーゼは,正常な記憶能を発揮する上で欠かすことができない.シアリダーゼ阻害剤(2,3-dehydro-2-deoxy-N-acetylneuraminic acid:DANA)をラット海馬に投与すると,モリス水迷路で評価した海馬依存性の空間記憶能が著しく低下する5).記憶能は,Neu4に対するsiRNAを脳室に持続投与することによっても低下する.また,DANAによって海馬苔状線維-CA3錐体細胞間やシャッファー側枝-CA1錐体細胞間におけるシナプス伝達効率の長期増強(long-term potentiation:LTP)が減弱することから,シアリダーゼはシナプス可塑性の制御に関わることが示されている5, 6).
新しいシナプスを形成するためには,神経接着因子(NCAM)のホモフィリック結合が必要である.NCAMにシアル酸重合体(PSA)が結合している場合,NCAMどうしの結合はシアル酸の負電荷によって抑制される.海馬の歯状回顆粒細胞は成人の脳でも新生されるが,新生直後の神経細胞はNCAMにPSAを発現している.この特性を利用して,PSA-NCAM抗体は新生神経のマーカーに利用されている.苔状線維の発達や発生時の神経回路形成において,新しいシナプスが作られるためにはNCAMからPSAが除去される必要がある7, 8).このことから,記憶形成時のシアル酸脱離は,PSA除去によるシナプス形成と関連している可能性がある.苔状線維はおよそ3か月ごとに新しい神経に入れ替わり,定期的に新しいシナプスを作り替えている.苔状線維終末にシアリダーゼ活性が比較的高いことからも9),苔状線維の新しいシナプス形成にシアリダーゼが関与することが示唆される.
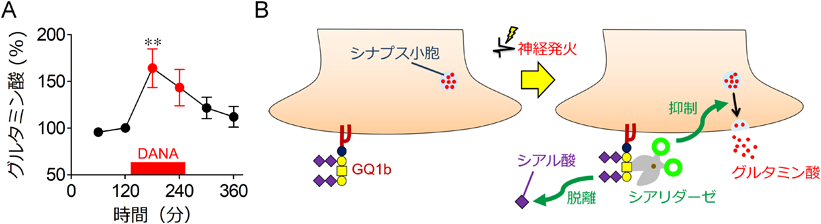
GQ1bなどのガングリオシドに結合したシアル酸は細胞内へのカルシウム流入や神経伝達物質の放出を促進することが報告されている10).このことから,神経発火時のシアル酸脱離は神経伝達物質の放出に影響することが推定された.そこで,シナプス開口放出を測定することができるスチリル蛍光色素FM1-43を利用して,海馬初代培養神経細胞のシナプス開口放出におけるシアリダーゼの役割を検討した11).神経の脱分極刺激や自発発火による開口放出は,シアリダーゼを阻害することによって促進された.また,海馬の主要な神経回路はグルタミン酸作動性神経によって構成される.ラットの海馬細胞外液に含まれるグルタミン酸量は,シアリダーゼ阻害剤であるDANAやDGNA(2,3-dehydro-2-deoxy-N-glycolylneuraminic acid)を作用させることによって増加した(図3A).シアリダーゼは興奮性神経伝達物質であるグルタミン酸の放出を抑制的に制御していると考えられる.シアリダーゼによる神経発火に伴ったシアル酸脱離は,グルタミン酸放出に対する負のフィードバック機構の一端を担うと考えられる(図3B).
ところで,ガングリオシドGQ1b/GT1aを認識する抗体(FS3抗体)を利用して免疫組織染色を行ったところ,DANAを作用させた後の海馬切片では染色強度が増加した.培養神経細胞の脱分極刺激による細胞内カルシウム濃度の増加もまた,DANAによって促進された.これらの知見から,シアリダーゼによるグルタミン酸の放出抑制には,ガングリオシドからのシアル酸脱離を介した細胞内へのカルシウム流入の抑制が関与することが示唆される.
哺乳動物が持つシアル酸の主要な分子種として,N-アセチルノイラミン酸(Neu5Ac)とN-グリコリルノイラミン酸(Neu5Gc)がある.Neu5Acは脳に豊富に存在し,記憶形成などの神経機能に重要な役割を担う.一方,Neu5GcはNeu5AcからCMP-Neu5Ac水酸化酵素(CMAH)などの作用を介して合成されるが,どの哺乳動物でも共通して脳においてはCMAHが発現していない.しかしながら,筆者らは以前にアイソトープラベルされたNeu5Gcを利用して,血中に存在するNeu5Gcが脳に移行することを見いだした12).興味深いことに,脳に移行するNeu5Gcは海馬に比較的高く集積する.Neu5GcはNeu5Acと比較してシアリダーゼによる加水分解を受けにくく,糖鎖から遊離されにくい2, 13).このようにNeu5Gcはシアリダーゼに抵抗を示すことから,シアリダーゼを介した神経機能の制御に影響を及ぼすと考えられる.実際に,脳にCMAHを強制発現させたトランスジェニックマウスでは,自発行動や物体認知記憶,髄鞘形成などに異常を示す14, 15).血中から脳に移行したNeu5Gcは,シアリダーゼの作用を抑制することによって神経機能に影響を及ぼすと考えられる.
本稿では,糖鎖構造が神経活動と連動して迅速に変化し,神経機能にフィードバック制御をかけることを解説した.現在,シアリダーゼの機能解析で得られた知見を基盤として,解析対象を中枢から末梢に,また,生理機能から疾患に広げている.今後は,糖鎖機能の解析ツール開発や解析対象をシアリダーゼ以外にも広げていきたい.
謝辞Acknowledgments
本稿で紹介したシアリダーゼ活性のイメージングプローブは,広島国際大学薬学部池田潔教授,および大坪忠宗准教授との共同研究の成果である.また,本稿で紹介した我々の研究成果は,当講座の高橋忠伸准教授,紅林佑希助教,会津大学短期大学部左一八教授,本学看護学部金澤寛明教授,愛知県がんセンター研究所疋田智也博士ならびに当講座の学生の方たちの協力なくしては得られなかったものである.ここに深く感謝申し上げます.
引用文献References
1) Monti, E. & Miyagi, T. (2015) Top. Curr. Chem., 366, 183–208.
2) Sumida, M., Hane, M., Yabe, U., Shimoda, Y., Pearce, O.M., Kiso, M., Miyagi, T., Sawada, M., Varki, A., Kitajima, K., & Sato, C. (2015) J. Biol. Chem., 290, 13202–13214.
3) Minami, A., Otsubo, T., Ieno, D., Ikeda, K., Kanazawa, H., Shimizu, K., Ohata, K., Yokochi, T., Horii, Y., Fukumoto, H., Taguchi, R., Takahashi, T., Oku, N., & Suzuki, T. (2014) PLoS ONE, 9, e81941.
4) Minami, A., Meguro, Y., Ishibashi, S., Ishii, A., Shiratori, M., Sai, S., Horii, Y., Shimizu, H., Fukumoto, H., Shinba, S., Taguchi, R., Takahashi, T., Otsubo, T., Ikeda, K., & Suzuki, T. (2017) J. Biol. Chem., 292, 5645–5654.
5) Minami, A., Saito, M., Mamada, S., Ieno, D., Hikita, T., Takahashi, T., Otsubo, T., Ikeda, K., & Suzuki, T. (2016) PLoS ONE, 11, e0165257.
6) Savotchenko, A., Romanov, A., Isaev, D., Maximyuk, O., Sydorenko, V., Holmes, G.L., & Isaeva, E. (2015) Neural Plast., 2015, 908190.
7) Seki, T. & Arai, Y. (1999) J. Comp. Neurol., 410, 115–125.
8) Di Cristo, G., Chattopadhyaya, B., Kuhlman, S.J., Fu, Y., Belanger, M.C., Wu, C.Z., Rutishauser, U., Maffei, L., & Huang, Z.J. (2007) Nat. Neurosci., 10, 1569–1577.
9) Minami, A., Shimizu, H., Meguro, Y., Shibata, N., Kanazawa, H., Ikeda, K., & Suzuki, T. (2011) Neuroimage, 58, 34–40.
10) Tanaka, Y., Waki, H., Kon, K., & Ando, S. (1997) Neuroreport, 8, 2203–2207.
11) Minami, A., Ishii, A., Shimba, S., Kano, T., Fujioka, E., Sai, S., Oshio, N., Ishibashi, S., Takahashi, T., Kurebayashi, Y., Kanazawa, H., Yuki, N., Otsubo, T., Ikeda, K., & Suzuki, T. (2017) J. Biochem., in press.
12) Taguchi, R., Minami, A., Matsuda, Y., Takahashi, T., Otsubo, T., Ikeda, K., & Suzuki, T. (2015) PLoS ONE, 10, e0131061.
13) Davies, L.R., Pearce, O.M., Tessier, M.B., Assar, S., Smutova, V., Pajunen, M., Sumida, M., Sato, C., Kitajima, K., Finne, J., Gagneux, P., Pshezhetsky, A., Woods, R., & Varki, A. (2012) J. Biol. Chem., 287, 28917–28931.
14) Naito-Matsui, Y., Davies, L.R., Takematsu, H., Chou, H.H., Tangvoranuntakul, P., Carlin, A.F., Verhagen, A., Heyser, C.J., Yoo, S.W., Choudhury, B., Paton, J.C., Paton, A.W., Varki, N.M., Schnaar, R.L., & Varki, A. (2017) J. Biol. Chem., 292, 2557–2570.
15) Collins, B.E., Fralich, T.J., Itonori, S., Ichikawa, Y., & Schnaar, R.L. (2000) Glycobiology, 10, 11–20.
著者紹介Author Profile
 南 彰(みなみ あきら)
南 彰(みなみ あきら)静岡県立大学大学院薬学研究院生化学講座講師.博士(薬学).
略歴1976年神奈川県横須賀市に生まれる.2000年静岡県立大学薬学部卒業.05年米国カリフォルニア大学バークレー校博士研究員.08年静岡県立大学薬学部助教.16年より現職.
研究テーマと抱負新しい糖鎖機能の分析ツールを構築することによって,これまでとは異なる視点から糖鎖の機能解明を試みる.今後は特に,認知機能における糖鎖の役割を明らかにしていきたい.
ウェブサイトhttp://w3pharm.u-shizuoka-ken.ac.jp/biochem/
趣味バイオリン,釣り.
 鈴木 隆(すずき たかし)
鈴木 隆(すずき たかし)静岡県立大学大学院薬学研究院生化学講座教授(薬学研究院長兼務).薬学博士.
略歴1956年静岡県に生まれる.79年静岡薬科大学卒業.84年同大学院博士課程修了.同年シノテスト研究所.91年静岡県立大学助手.94年同講師.96年同助教授.97~99年米国St. Jude Children’s Research Hospital客員研究員.2006年より静岡県立大学教授.
研究テーマと抱負私たちは糖鎖が関わるすべての生命現象に興味を持っています.特に,ウイルス感染症,中枢神経機能,癌における役割を分子レベル,遺伝子レベルで解明しています.さらに,その成果を応用することを目指しています.
ウェブサイトhttp://w3pharm.u-shizuoka-ken.ac.jp/biochem/
趣味野菜栽培,釣り.