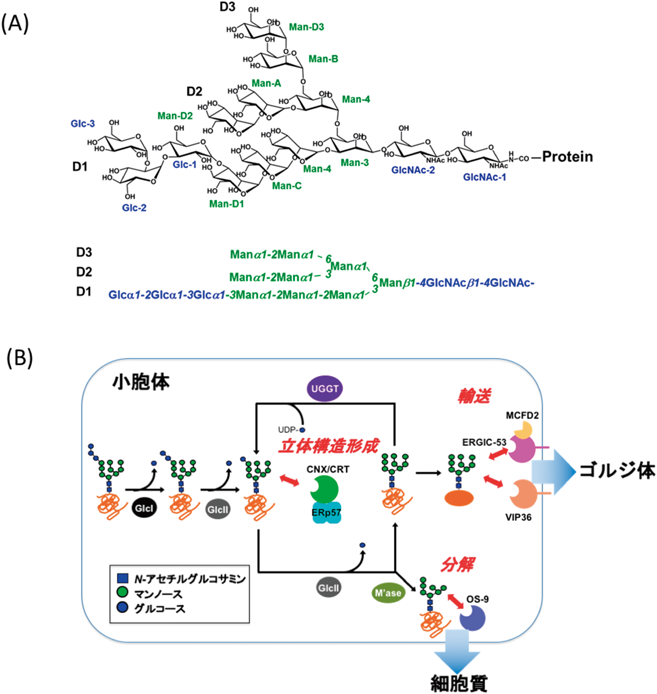
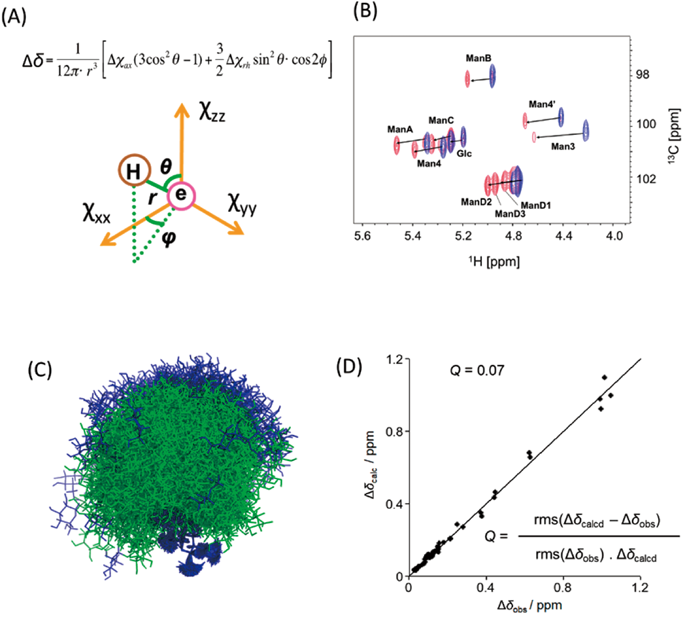
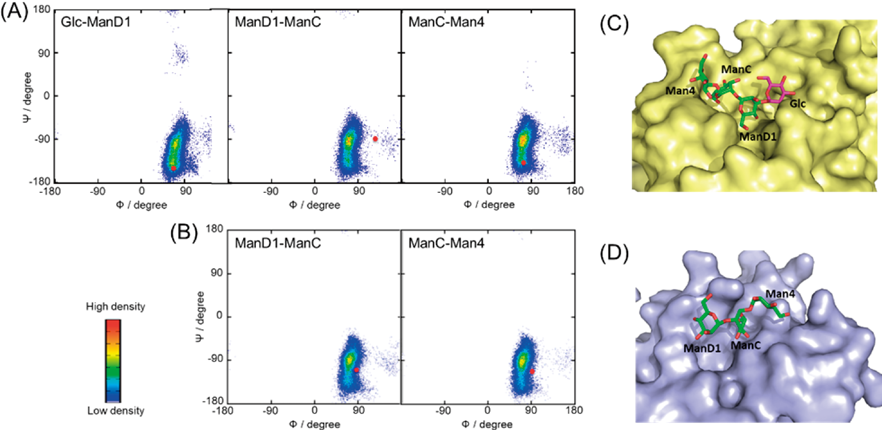
常磁性NMR法と計算科学を組み合わせた糖鎖の動的コンホメーション解析Dynamic conformation analysis of oligosaccharides by combining paramagnetism-assisted NMR spectroscopy and molecular dynamics simulation
1 名古屋市立大学・大学院薬学研究科Graduate School of Pharmaceutical Sciences, Nagoya City University ◇ 〒467–8603 名古屋市瑞穂区田辺通3–1 ◇ 3–1 Tanabe-dori, Mizuho-ku, Nagoya 467–8603, Japan
2 自然科学研究機構・生命創成探究センターExploratory Research Center on Life and Living Systems, National Institutes of Natural Sciences ◇ 〒444–8787 岡崎市明大寺町東山5–1 ◇ 5–1 Higashiyama, Myodaiji, Okazaki, Aichi 444–8787, Japan
発行日:2018年4月25日Published: April 25, 2018