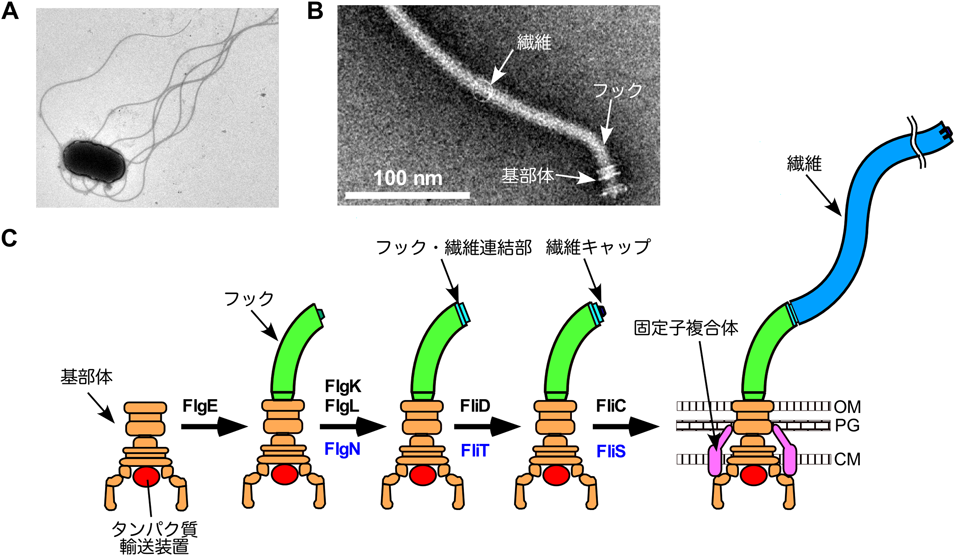
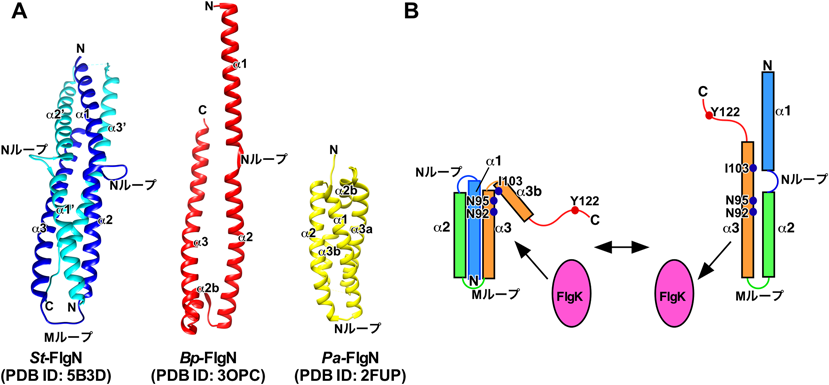
細菌べん毛特異的輸送シャペロンFlgNの構造と機能Structure and function of a bacterial flagellar export chaperone FlgN
大阪大学大学院生命機能研究科Graduate school of Frontier Biosciences, Osaka University ◇ 〒565–0871 大阪府吹田市山田丘1–3 ◇ 1–3 Yamadaoka, Suita, Osaka 565–0871, Japan
発行日:2018年4月25日Published: April 25, 2018