生物体内に極微量存在する鉄(Fe),亜鉛(Zn),銅(Cu)等のいくつかの金属元素は,生物が生きていく上で必須であることはよく知られている.これらの金属元素(生体金属と呼ぶ)は,金属特有の化学的な特性,すなわち配位結合,酸化還元,ルイス酸などの特性を活用して,タンパク質などの有機物のみでは絶対にできない生理反応に関与している.たとえば,気体小分子の担体,特異な構造の構造因子あるいは酵素の活性中心として機能しており,さまざまな生理現象において中心的な役割を果たしている.生物無機化学は,これら生体金属の役割を分子・原子・電子のレベルで解き明かしていく研究分野であり,構造生物学の手法との共同研究により,金属結合酵素や金属結合タンパク質の分子構造を基盤にした理解の進展に大きく寄与してきた.しかし,今後のこの分野の深化を考えると,以下のような疑問にも応えていかねばならない.すなわち,金属ならびに金属酵素・金属タンパク質は,細胞内ではどのような構造をとり,どのように機能しているのであろうか.単離精製した金属酵素・金属タンパク質の研究から得られた知見は,そのまま細胞内でのそれらの機能理解に拡張できるのであろうか.また,単離精製した金属酵素・金属タンパク質でも,止まった構造で機能しているのでなく,常に構造的な揺動(fluctuation)があり,また構造変化を伴って機能しているはずである.さらには,酵素反応時に必ず現れる短寿命反応中間体の情報は機能理解に必須である.以上のような疑問に対しては,静的構造だけではなく,動的構造(dynamic structure)あるいは構造ダイナミクス(structural dynamics)の情報を基盤に答えねばならないであろう.本稿前半では,鉄含有酵素である一酸化窒素還元酵素を対象として構造ダイナミクスを基盤にした機能解析の研究を紹介する.
一方,生体は生体金属を必ず外部から取り込まれねばならず,その際の吸収,運搬,濃度感知,貯蔵など(金属の生体内動態あるいは生体内金属動態)がどのように行われているのかは非常に興味深い.この生体内金属動態には数々のタンパク質が関与しており,生体内の金属濃度を厳密に制御している.生体内金属動態の破綻は,生体金属の過剰や不足を招き,疾病の原因になる.また,生体に必要ない金属の取り込みも疾病の原因になる.イタイイタイ病のカドミウム(Cd)や水俣病の水銀(Hg)などはその例である.本稿後半では,病原菌のヒトへの感染増殖に必須の鉄取り込みに着目し,その構造ダイナミクス研究の例と将来展望を述べる.
1)脱窒
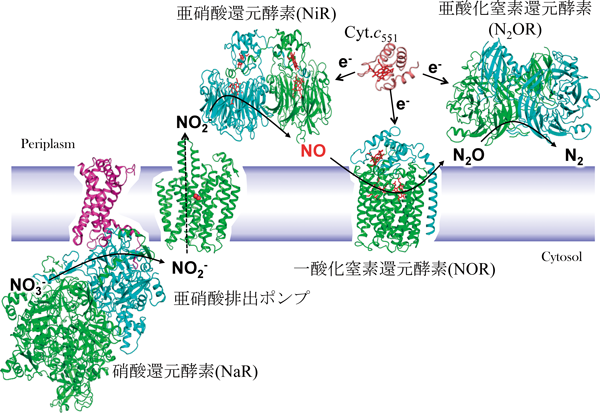
一酸化窒素還元酵素(nitric oxide reductase:NOR)は,微生物の嫌気呼吸の一種である脱窒(denitrification)に関与する酵素である.脱窒は,硝酸イオン(NO3−),亜硝酸イオン(NO2−)が一酸化窒素(NO),亜酸化窒素(N2O)の反応中間物質を経て,最終的に窒素分子(N2)まで順次還元される過程であり(NO3−→NO2−→NO→N2O→N2),地球上の窒素循環において重要な役割を果たしている.脱窒の4段階の反応には,硝酸還元酵素(nitrate reductase:NaR, Mo/Fe含有),亜硝酸窒素還元酵素(nitrite reductase:NiR, CuあるいはFe含有),一酸化窒素還元酵素(NOR, Fe含有),亜酸化窒素還元酵素(nitrous oxide reductase:N2OR, Cu含有)の4種類の金属酵素が各々の反応を触媒している.すべての酵素の結晶構造は明らかにされている(図1).
脱窒において一酸化窒素NOはNiRによって次式のように産生されるが, ラジカル分子であり反応性が非常に高く,多くの生体物質と反応してそれらの機能を阻害する非常に細胞毒性の高い分子である.しかし,脱窒微生物においては,脱窒条件下での生育ではNOは検出できない.このことは,NiRによって産生されたNOが細胞内外に拡散することなく速やかに還元・無毒化されていることを示しており,それを担っているのがNORである. NORには脱窒カビの酵素と脱窒菌の酵素があり,それらの構造は我々のグループが明らかにしている1–3).NiRによって産生されたNOはいかにNORに受け渡されるのか? NORはいかにNOを無毒化しているのか? この2点は構造ダイナミクスにより解き明かさねばならない問題であった.
2)NiR-NOR複合体
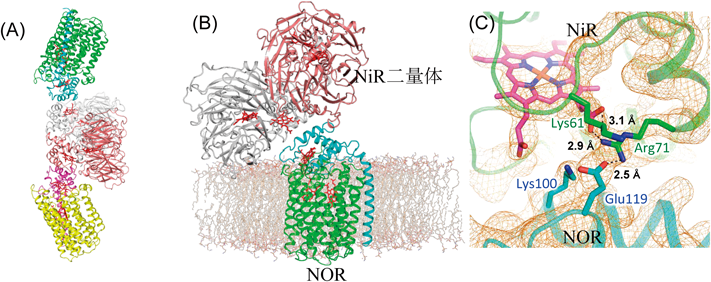
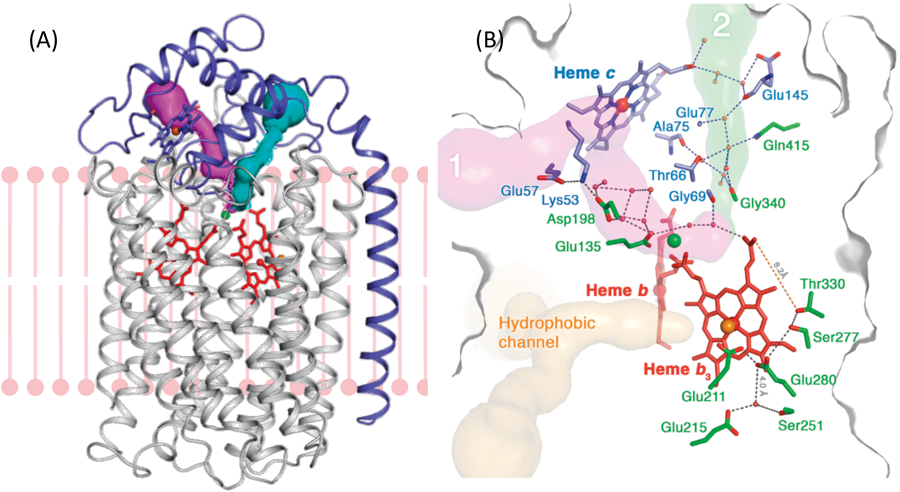
脱窒菌のNiRは水溶性酵素であり,一方のNORは膜結合性酵素である.NiRからNORへのNOの受け渡しには,あるタンパク質が仲介するか,直接受け渡すかの2通りの方法が考えられた.後者の可能性を検討する目的で,NiRとNORの共結晶を作製し,X線結晶構造解析でその構造を決定した4).その結果,NiR二量体に2分子のNORが相互作用した構造が得られた(図2A).この構造から,実際の細胞中では膜に固定されたNOR 1分子にNiR二量体が相互作用していると推定した(図2B).細胞膜を含むNiR-NOR複合体に,分子動力学(MD)シミュレーションを適応したところ,この複合体は十分に安定であることが確かめられ,このような複合体構造が細胞中でも安定に存在する可能性を支持できた.実際,両酵素の接触界面で塩橋を作っている残基(NORのLys100とGlu119)に変異を導入したところ(図2C),その変異体を含む脱窒菌(緑膿菌)は硝酸塩存在下の嫌気条件(脱窒条件)で培養しても生育しなかった.さらにNiRにより産生されたNOの挙動をMDでシミュレーションした.NOは疎水性分子であるため,水中で産生されても速やかに細胞膜内に移動し,その近くに存在するNORの疎水性チャネルを通過して,拡散律速に近い速度(10−8 s−1)で酵素反応中心まで到達した.実験的にもこの迅速なNO結合は確かめられている.すなわち,NiR-NOR複合体形成は,水溶性タンパク質NiRを細胞膜と膜タンパク質NORの近傍に位置させることにより,産生されたNOを細胞内に拡散させずに速やかに消去するシステムと結論した.
以上の研究は,細胞中のNiRとNORを直接観測したものではないが,細胞内での複合体形成の可能性を強く示唆している.最近,脱窒菌内で脱窒酵素・タンパク質群(NaR, NiR, NOR, N2OR,電子供与体,トランスポーター)が超分子複合体を形成している可能性も指摘されている5).また,クライオ電子顕微鏡を用いた研究により,ミトコンドリアの好気呼吸酵素群の超分子複合体構造が報告されている6).細胞内において,一つの酵素の生成物が他の酵素の基質としてスムーズに受け渡されていく構造的な基盤が示され始めている.
3)NO還元反応の構造ダイナミクス
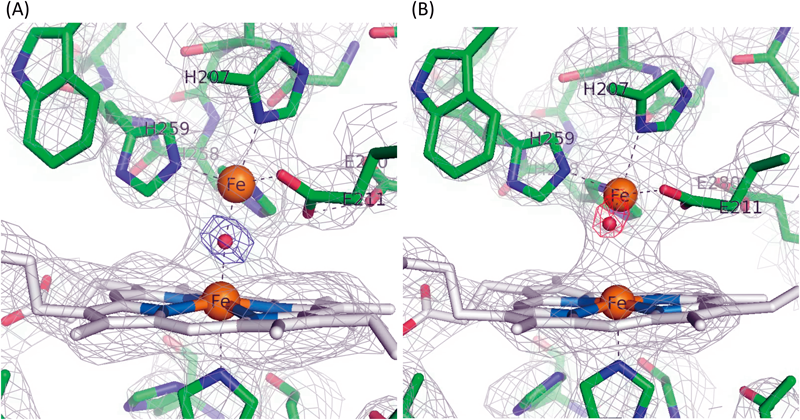
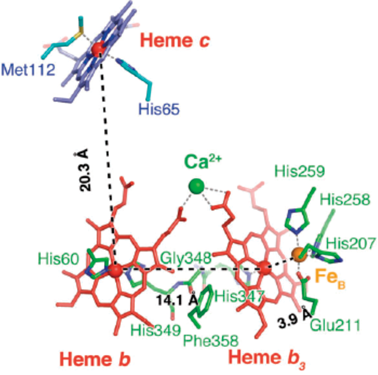
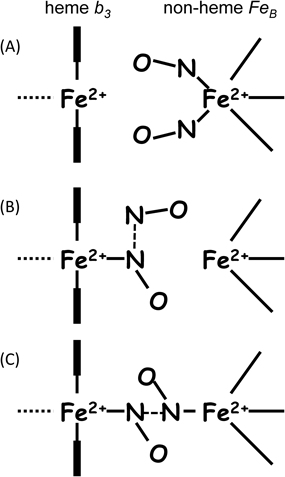
NORによるNO還元反応においては,2プロトン(H+)と2電子(e−)によってN–N結合の生成とN–O結合の開裂が行われる.この反応を構造ダイナミクスの観点から解き明かした.脱窒菌のNORの活性中心は,ヘム鉄(heme b3)と非ヘム鉄(FeB)からなる複核中心である(図3A).酵素反応時に,この複核活性中心へは,電子供与体であるチトクロムc551あるいはアズリンからの電子が,NOR内に存在するヘムc(heme c)と低スピン状態のヘムb(heme b)を経て,複核中心(heme b3とFeB)に供与される(図4).結果的に複核中心の二つの鉄は三価Fe3+から二価Fe2+に還元される(図3B)7).この還元型NORに,NiRにより産生されたNOが配位し酵素反応が開始する.NOは鉄に結合してから数ミリ秒の短寿命反応中間体の構造が重要である.この反応中間体については,完全還元(Fe2+)型の複核中心へ2分子のNOが配位した構造と考えられ,今までに3種類の構造が提案されている(図5)が,まだ確定には至っていない(後述).しかし,いずれの反応中間体でも,この後にはおそらく次亜硝酸(hyponitrite:−O–N=N–O−)が生成し(N=N結合生成),プロトン(H+)供与によってN–O結合が開裂し,N2OとH2Oが産生すると考えられている.
N–O結合開裂に必要な触媒プロトンの供給経路に関しては,結晶構造とMDシミュレーションが重要な知見を与えた.酵素の細胞質側が疎水性の残基で形成されているのに対して,ペリプラズム側に水層から活性中心へとつながる二つの水素結合ネットワークが見いだされた(図6).MDシミュレーションによれば,二つのうちGlu57を入り口としAsp198を経るネットワークがプロトン供給経路と決定できた8).実際,NORのE57AとD198N変異体は,NO還元活性を喪失した.
4)短寿命反応中間体の配位・電子構造解析の手法開発
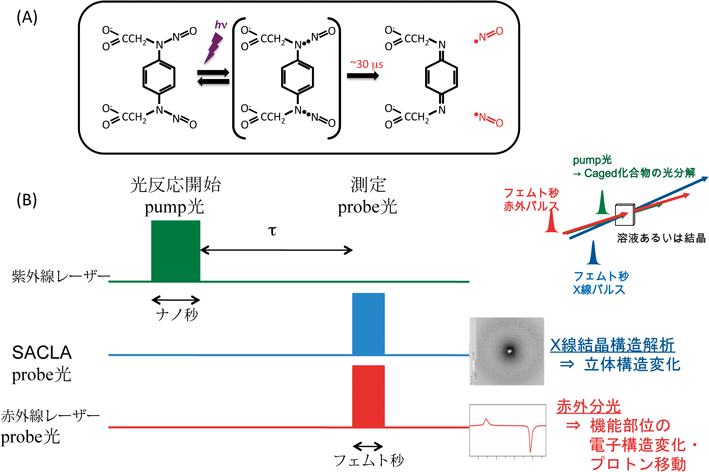
NORの酵素反応を構造ダイナミクス基盤で理解していく上では,先述の短寿命反応中間体の情報は欠かせない.反応中間体のNOの配位構造とそのNO結合型酵素の電子構造を明らかにする手法としては,時分割測定が必須であろう.特に,時分割X線結晶構造解析法と時分割赤外分光法が有力な手法である.これらの手法では,紫外光を照射すると数十マイクロ秒の時間でほぼ定量的にNOを発生するcaged NOと呼ばれる化合物を反応開始に用いる.すなわち,NORとcaged NOを含む試料に紫外光(pump光)をパルス照射してNO結合反応を開始し,反応中間体が生成するまでの時間(遅延時間)を待って,配位構造や電子状態をモニターする光(probe光)をパルス照射する手法である(pump-probe法).時分割赤外分光法ではprobe光としてフェムト秒の赤外線レーザーを光源とし,時分割X線結晶構造解析法ではフェムト秒のX線レーザーを光源とする(図7).短時間のパルスでも十分に明るい光源が開発されたことが,このような測定が可能となった一番の要因である.特にX線自由電子レーザー施設SACLAによるところが大きい.時分割X線結晶構造解析法に必要な多量な微結晶を得ることができたので,脱窒カビNORにこれらの手法を適用した結果を以下に示す9).
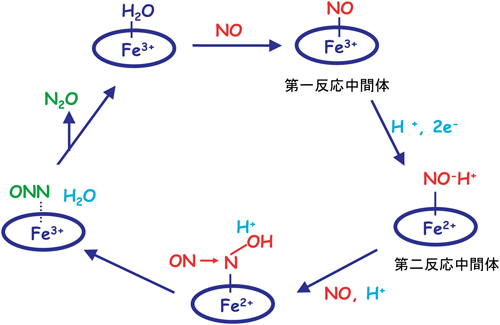
脱窒菌と脱窒カビのNORは,同じ反応(式1)を触媒するが,その構造と性質はまったく異なる.脱窒カビのNORは水溶性酵素であり,ヘムbを1分子のみ活性中心に含み,その反応機構は図8のように提案されている10).この機構の特徴は,Fe3+にNOが結合した状態が第一反応中間体である.このFe3+–NO状態が近傍に存在するNAD(P)Hにより直接2電子還元[H+の移動も含まれるので,ヒドリド(H−)移動]され,第二反応中間体[Fe3+–NO]2−H+が生成する.第二反応中間体は電子過剰の状態であるので,もう1分子のNOと反応しやすく,この反応によってN–N結合ができあがる.溶媒に水から水素結合ネットワークによって運ばれたH+によって,N–O結合が開裂する.時分割X線結晶構造解析法と時分割赤外分光法を適用した結果,第一反応中間体ではFe–N–O構造において,Fe–N距離は1.67 Å,N–O距離は1.15 Å,N–O伸縮振動は1853 cm−1,F–N–O角度は185°と求まった.第二反応中間体では,Fe–N距離は1.81 Å,N–O距離は1.25 Å,N–O伸縮振動は1333 cm−1,F–N–O角度は126°と求まり,第一反応中間体の2電子還元型の構造と推定した.
以上のように,caged NOと時分割X線結晶構造解析法あるいは時分割赤外分光法を組み合わせた手法は,短寿命反応中間体の配位構造・電子構造を議論する上で非常に有力である.今後は,脱窒菌NORに本方法を適用し,その酵素反応の分子機構を確立したい.
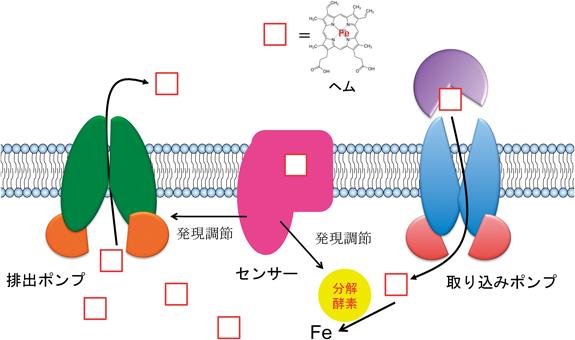
緑膿菌,黄色ブドウ球菌,ペスト菌,ジフテリア菌,炭疽菌などの病原菌は,感染先の赤血球ヘモグロビンを鉄源としている.図9に示したように,これら病原菌は自身の細胞膜に存在するヘムインポーター(ヘム取り込みポンプ)を使って,ヘモグロビンのヘムを取り込む.ヘムインポーターはATPの加水分解エネルギーを使ってヘムを能動的に膜輸送するABCトランスポーターに属している.ヘム輸送に際して,病原菌はヘム濃度センサータンパク質によってヘム濃度を感知し,取り込んだヘムを分解するヘム分解酵素(ヘムオキシゲナーゼ)を発現させる.ヘムオキシゲナーゼは酸素添加反応によってヘム(鉄−ポルフィリン錯体)のポルフィリン部分を分解して鉄を取り出し,それを増殖に使う.一方,病原菌がヘムを取り込みすぎると,過剰のヘムは酸素分子と反応し活性酸素種を発生させるので,病原菌にとって致死的になってしまう.そのため,病原菌のヘム濃度センサータンパク質は,やはりABCトランスポーターに属するヘムエクスポーター(ヘム排出ポンプ)を作動させて,過剰なヘムを細胞外に排出する.ヘムのインポーター,エクスポーター,濃度センサー,分解酵素のどれが機能しなくなっても,病原菌は増殖できないので,これらの阻害剤はよい抗菌薬になる.これらタンパク質の静的構造だけでなく構造ダイナミクスを明らかにできれば,病原菌の鉄動態の機構を分子レベルで理解できるだけでなく,阻害剤の設計にもつながる.本節では,ヘムインポーターに関する我々の研究成果を述べる11).
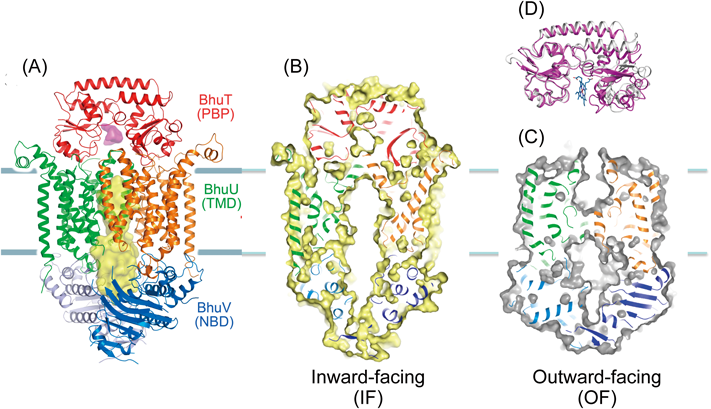
院内感染菌(Burkholderia cenocepacia)のヘムインポーターは,細胞のペリプラズムにおいてヘムを運搬するタンパク質(BhuT),細胞膜でヘムを取り込む膜貫通領域(BhuU),ATP加水分解領域(BhuV)の3種類のサブユニットが,BhuT 1分子,BhuU 2分子,BhuV 2分子からなるヘテロ五量体構造である(BhuTUV,図10A).我々の構造には,基質であるヘムとATPの両方とも結合していなかった.
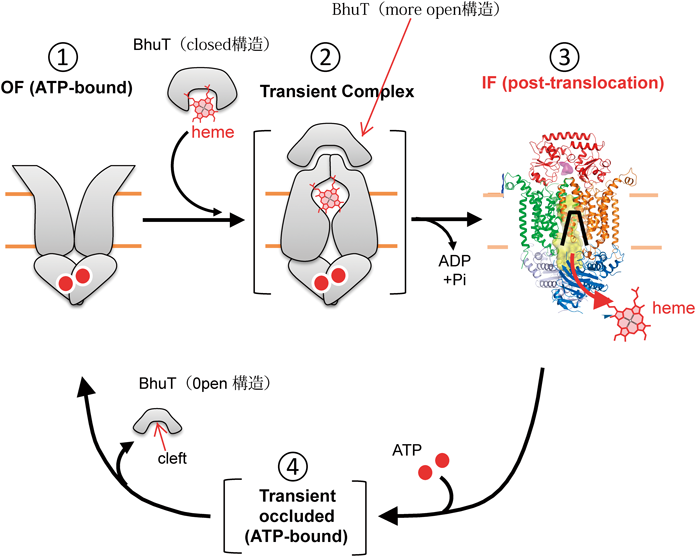
BhuU 2分子の接触界面にはヘムの通過チャネルが存在し,我々はこのチャネルが細胞内に開いた(IF:inward-facing)構造を決定できた(図10B).これはヘムを取り込んだ後の構造と考えられた.一方,スイスのグループはペスト菌のヘムインポーターの構造を決定しており,ヘム運搬タンパク質は結合しておらず,チャネルは細胞外に開いた(OF:outward-facing)構造であった(図10C).ヘムを運ぶ前の構造と考えられる.一方,ヘム運搬タンパク質BhuT単独で,ヘムが結合していない構造と結合した構造の2種類を解いている(図10D).以上の構造情報を基盤に,ヘムインポーターによるヘム膜輸送の機構を以下のように提案した(図11).
- BhuTにはヘムが結合できる間隙(cleft)が存在し,その間隙はヘムがないときには閉じた(closed)構造をとり,ヘム結合により開いた(open)構造へと変換される(図11①).
- ヘム結合BhuTは,OF構造のBhuUに静電的相互作用によって結合する(図11②).
- しかし,BhuTは間隙がさらに開いた(more open)構造をとらないと,相互作用はできず結合もできない.この構造により,ヘムとBhuT間隙との相互作用は弱まり,ヘムはBhuUの疎水性チャネルに移動する.
- BhuVに結合していたATPの加水分解により,BhuUはOFからIF構造へと変化する(図11③).
- IF構造のBhuUのチャネルは疎水的であるが,OF構造では負電荷を持ったアミノ酸が露出してくる.この負電荷とヘムのプロピオン酸の負電荷の反発により,ヘムは細胞内へ放出される(図11③).
- ATPがBhuVに結合し(図11④),BhuUはIFからOF構造に,BhuTはBhuUから解離し,元の状態に戻る.
以上の病原菌のヘム膜輸送機構は,あくまでも静的な構造情報のつなぎ合わせである.現在はMDシミュレーションを行うと同時に,先に述べたcaged化合物(この場合はcaged ATP)とXFELを組み合わせた動的な構造解析を計画しており,構造ダイナミクスを基盤に理解を拡げていきたい.同時にヘムのエクスポーターやヘム濃度センサーへもこの手法を拡張していく.